| Review article |
Open Access |
|
| Hiroshi Ohrui* |
| Yokohama College of Pharmacy, 601 Matanocho, Totsukaku, Yokohama, Japan |
| *Corresponding authors: |
Hiroshi Ohrui
Yokohama College of Pharmacy
601 Matanocho, Totsukaku
Yokohama, Japan
Email: h.ohrui@hamayaku.ac.jp |
|
| |
| Received April 26, 2012; Published August 28, 2012 |
| |
| Citation:Ohrui H (2012) 4’-C-Ethynyl-2-Fluoro-2’-Deoxyadenosine (EFdA): A Two Site Modified 2’-Deoxyadenosine Derivative which Could Prevent the Emergence of Resistant HIV Mutant and has Supremely High Anti-HIV Activity and Low Toxicity 1:281. doi:10.4172/scientificreports.281 |
| |
| Copyright: © 2012 Ohrui H, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. |
| |
| Abstract |
| |
| The development of 2’-deoxy-4’-C-ethynyl-2-fluoroadenosine (EFdA,) which has supremely high anti-HIV activity and low toxicity and prevents the emergence of resistant HIV mutants was achieved by the studies based on the author’s working hypotheses, and a very important discovery that the substrate selectivity of reverse transcriptase of HIV (RNA-dependent DNA polymerase) is different from that of human DNA-dependent DNA polymerases was made in this studies. |
| |
| Keywords |
| |
| 4’-C-substituted-2’-deoxynucleoide; Decrease of toxicity of nucleoside; Nucleoside reverse transcriptase inhibitor (RT); Prevention of the emergence of resistant HIV mutant; Different substrate selectivity between RT and human DNA polymerases |
| |
| Introduction |
| |
| Many emerging viral infectious diseases, for example, the spreads of AIDS, Flu, West Nile Virus, SARS, and so on, are causing major threats to global public health. Therefore, the development of highly active and lowly toxic antiviral drugs is required. |
| |
| Since the discovery of 3’-C-azido-3’-deoxythymidine (AZT) as an anti-human immunodeficiency virus (anti-HIV) agent by Dr. Hiroaki Mitsuya in 1985, [1] many 2’,3’-dideoxy nucleoside (ddN) analogs have been developed as nucleoside reverse transcriptase inhibitor (NRTI). However, resistant HIV mutants against all these NRTI have emerged very rapidly and easily. |
| |
| Highly Active Antiretroviral Therapy (HAART) using two or more NRTI and protease inhibitors has dramatically improved the quality of life and prognosis of patients infected by HIV [2,3]. However, the existing HAART has several critical problems that remain to be solved. These problems are: (i) the emergence of new drug-resistant HIV mutants, (ii) drug side effects, and (iii) need to take large dosages of drugs. Therefore, the development of new anti-HIV drugs that solve these problems is urgently needed. |
| |
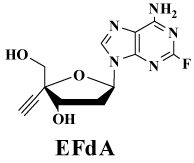
| This review describes the working hypotheses for the development of modified nucleosides which could solve the above problems, the results of the validation studies of the hypotheses, the perspective of the most promising anti-HIV active 2’-deoxy-4’-C-ethynyl-2- fluoroadenosine (EFdA) (Figure 1) developed by the studies, the mechanism of inhibition of reverse transcriptase (RT) and DNA polymerases by EFdA, an improved synthesis of EFdA, a discovery of that substrate selectivity is different between RT and and human DNA polymerases which should be very important for the development of excellent anti-HIV modified nucleosides. |
| |
|
|
Figure 1: Working hypotheses for the development of modified nucleosides which could solve the problems of the existing HAART. |
|
| |
| The way to prevent the emergence of resistant HIV mutants against modified nucleoside. Design of 4’-C-substituted-2’-deoxynucleoside (4’SdN) as the modified nucleoside which could prevent the emergence of resistant HIV mutants |
| |
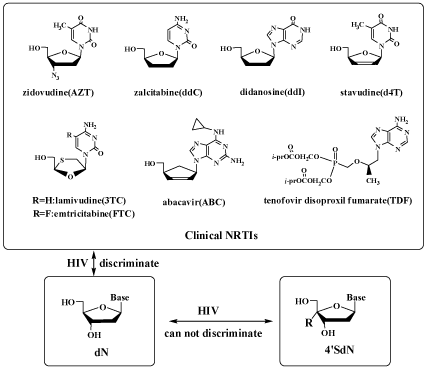
| The structures of the clinical NRTI are shown in Figure 2. |
| |
|
|
Figure 2: The structures of the clinical NRTI. |
|
| |
| All the NRTI belong to the family of 2’, 3’-dideoxy nucleoside (ddN). The ddN structure (which does not have 3’-OH) has been long assumed essential for the nucleoside derivatives to be the chain terminator of RT. However, HIV mutants resistant to all the clinical NRTI have emerged very rapidly and easily. |
| |
| I speculated that the emergence of HIV mutants resistant to ddN indicated that the resistant HIV mutants acquired the ability to differentiate the ddN from physiologic 2’-deoxy nucleoside (dN) and do not accept the ddN into the active center of RT and/or cut off the incorporated ddN from the proviral DNA terminus, and therefore, the anti-HIV nucleosides that could prevent the emergence of drugresistant HIV mutants must satisfy the following two conditions. |
| |
| 1. To prevent the emergence of resistant HIV mutants, the modified nucleosides must prevent the differentiation by HIV from physiologic dN. Since the significant difference of the structure between ddN and physiologic dN is whether they have 3’-OH, the modified nucleosides that could prevent the emergence of resistant HIV mutants must have 3’-OH. |
| |
| 2. In spite of having 3’-OH, the modified nucleoside must be the chain terminator of RT- catalyzed biosynthesis of proviral DNA. |
| |
| Based on the following five hypotheses, 2’-deoxy-4’-C-substituted nucleoside (4’SdN, Figure 2) was designed as a nucleoside that could satisfy the above two conditions: |
| |
| ââ¡ It would be difficult for HIV to differentiate 4’SdN from dN because 4’SdN has all the functional groups of dN. |
| |
| ââ¡ The introduction of a substituent at 4’-position makes the 3’-OH into a very unreactive neopentyl-type secondary alcohol. Thus, the 3’-OH of 4’SdN will be used for HIV to mistake 4’SdN for dN, but is too unreactive to be used for the elongation of proviral DNA by RT. Therefore, 4’SdN could be NRTI. |
| |
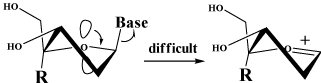
| ââ¡ The steric hindrance between 3’-OH and 4’-substituent changes the conformation of the furanose ring of 4’SdN preferably to the 3’-endo conformation (N-type) [4]. This results in 4’SdN being less susceptible to both enzymatic and acidic glycolysis than dN and ddN, because in glycolysis, the oxygen atom of the furanose ring participates to form a coplanar oxocarbonium, however, the conformational change makes the oxygen atom of 4’SdN difficult to participate to form a coplanar oxocarboium ion (Figure 3). |
| |
|
|
| |
| ââ¡ Further, the electron-withdrawing 3’-OH makes 4’SdN more stable to acid than is ddN even with purines. Thus, various purine derivatives can be made in this way |
| |
| ââ¡ The lipophilic substituent at the 4’-position imparts more lipophilicity to 4’SdN, thus enabling them to penetrate the cell membrane efficiently. This possibly enhances their bioavailability. |
| |
| The way to decrease the toxicity of nucleosides |
| |
| If DNA polymerases also mistake 4’SdN for dN, 4’SdN should be highly toxic. |
| |
| The clinical NRTI drugs are also the chain terminators of DNA polymerases and therefore toxic and have fairy severe side effects. However, the ddN have been used as anti-HIV drugs, meaning that human DNA polymerases accept ddN less than RT does, thus, the substrate selectivity between RT and human DNA polymerases must be different. |
| |
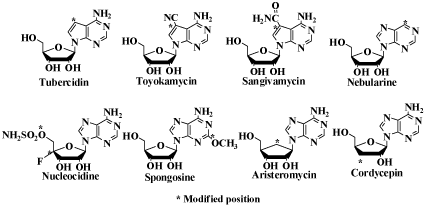
| The structures of the representative nucleoside antibiotics [5] are shown in Figure 4. |
| |
|
|
Figure 4: Representative nucleoside antibiotics. |
|
| |
| Most of them are nucleoside derivatives modified at one site of the physiological nucleosides. Though they are highly anti-microbial active, they are highly toxic, too. Therefore, they can not be clinically used. |
| |
| In the 1960s and 1970s, many organic chemists modified these antibiotics expecting to get nucleoside derivatives with better biological activity. However, the additionally modified nucleosides decreased or lost the antimicrobial activity. The same results were obtained with synthetic modified nucleosides. The one site modified nucleosides were highly antimicrobial active and highly toxic, and the additional modification of these nucleosides resulted in the decrease or loss of the activity. These results indicated that the intracellular enzymes do not or hardly recognize the nucleoside derivatives modified at two or more sites of physiologic nucleosides as their substrates. Thus, there is a chance of decreasing the toxicity of 4’SdN further by additional modification. |
| |
| Examination of the validity of the working hypotheses with 4’-C-methyl nucleosides |
| |
| At first, in order to examine the validity of the working hypotheses, 4’-C-methyl-D-ribofuranosyl nucleoside (MN), 4’-C-methyl-2’- deoxy-D-ribofuranosyl nucleoside (MdN), 4’-C-methyl-2’,3’- dideoxy-D- pentofuranosyl nucleoside (MddN) and 4’-C-methyl-2’,3’- didehydrodideoxy-D- pentofuranosyl nucleoside (Md4N) (Tale 1) were synthesized and evaluated for their biological activity [6]. |
| |
|
|
Table1: anti-HIVactivityof4'-C-methyl-2'-deoxynucleosides |
|
| |
| MdN showed remarkable biological activity (both anti-HIV activity and toxicity), but MddN and Md4N did not show notable biological activity. |
| |
| MN did not show any anti-HIV activity and toxicity at all. |
| |
| These results indicated the importance of the 3’-OH for biological activity. Further, we demonstrated that 5’-O-triphosphate of both 4’-C-methyl-2’- deoxycytidine (MdCTP) and 4’-C-methyl-Darabinofuranosyl cytosine (MAraCTP) are the chain terminators of calf thymus DNA polymerase α and recombinant rat DNA polymerase β [7]. These results indicated that 4’SdN is NRTI, although further study of MdCTP with RT was not performed. |
| |
| They were synthesized by glycosidation of 1,2-di-O-acetyl-3,5- di-O- benzyl-4-C-methyl-D-ribo-furanose [8] which was prepared from D-glucose through 3-O-benzyl-4-C-hydroxymethyl-1,2-Oisopropylidene- α-D-ribo-furanose [9], and nucleobases and then deoxygenation of the 2’-hydroxyl groups of MN. |
| |
| It should be noted that the 4’-C-methyl ribofuranose derivatives took longer reaction time to complete the glycosidation reaction with silylated nucleoside bases than that of normal ribofuranose derivatives, indicating the lower reactivity of the 4’-C-methyl ribofuranoses than normal ribofuranoses. In addition, the 3J1’,2’ values of MN were larger (4~7Hz) than those of the normal β-ribonucleoside (0~1Hz), indicating the change of the conformation of the furanose ring of the 4’-C-substituted-β-D-ribofuranosyl nucleosides [8]. |
| |
| Structure-Activity Relationship (SAR) of 4’SdN |
| |
| Next, to study the SAR of 4’SdN and develop 4’SdN having more potent anti-HIV activity and less toxicity than MdN, 4’SdN having various kinds of 4’-C-substituents and nucleobases were synthesized and evaluated for their biological activity [10-17]. While we were working on our project, the anti-HIV activity of several 4’SdN was reported by the Syntex group [18-23] and others [24-25]. Therefore, the anti-HIV activities of 4’SdN that we studied together with those reported by other groups are listed in Table 2. |
| |
|
|
Table2: Anti-HIVactivityof4'-C-substituited-2'-deoxynucleosides. |
|
| |
| The SAR of 4’-C-substitued nucleosides against HIV are summarized as follows: |
| |
| 1. The estimated relative order of anti-HIV activity is as follows: |
| |
| C≡CH > CN > N3 > CH=CH2 > Me= Et > C≡C-CH3. Interestingly, the order is the reverse of the –ΔG° values between equatorial and axial substituents on a cyclohexane ring: CN < C≡CH < CH=CH2 < Me < Et < tBu. Thus, these results indicate that the sterically less demanding substituent at the 4’-position gives more potent anti-HIV activity, thus, the 4’SdN resembling more closely to the physiologic nucleosides has higher anti-HIV activity. |
| |
| 2. Purine analogs are generally less toxic than pyrimidine. Although 2’-deoxy-4’-C-ethynyl-5-fluorocytidine (EFdC), which is a nucleoside derivative modified at two positions of physiologic 2'-deoycytidine, gave a very acceptable Selectivity Index (SI=CC50/ EC50) with MT-4 cells, it was toxic with other cells (Kohgo, Yamasa Corporation, private communication ). Therefore, we did not study on EFdC any further. |
| |
| 3. Arabino analogs are less active and less toxic compared with their corresponding 2’-deoxy counterparts. |
| |
| 4. 4’SddN did not show anti-HIV activity. |
| |
| 5. The L-isomers of 4’SdN have no anti-HIV activity,15) although it is known that the L-enantiomer of 2’,3’-dideoxy-3’-thiacytidine (3TC) is as active as the D-enantiomer and less toxic than the D-isomer [25]. This may be due to that the L-isomers of 4’SdN are too much modified to be recognized by RT as its substrates. |
| |
| Synthesis of purine derivatives of 2’-deoxy-4’-C-cyanonucleoside (CNdN) and 2’-deoxy 4’-C-ethynylnucleoside (EdNs) and their anti- HIV activity |
| |
| The mentioned results led us to study the biological activity of purine derivatives of CNdN and EdN [26]. Although 4’SdN in our project had been synthesized by the glycosidation of 4-C-substituted- D-ribo-furanose derivatives and nucleobases, this synthetic route incurred some problems, as follows: |
| |
| 1. Preparation of 4-C-substituted-D-ribofuranoses and their conversion to the desired 4’SdN require multistep reactions, and their total yields are low. |
| |
| 2. 4-C-Substituited-D-ribofuranose derivatives have low reactivity in glycosidation reactions, especially when the substituent is an electron-withdrawing group such as a cyano group. |
| |
| (In contrast, the low reactivity of the anomeric position of 4-C-substituted furanose derivatives indicates that 4’SdN will be more stable to acidic hydrolysis than dN, ddN, and d4N will.) |
| |
| These problems prompted us to develop another method of preparing purine derivatives of 4’SdN, which starts from dN [26]. |
| |
| The biological activities of the purine derivatives of CNdN and EdN are summarized as follows (Table 3): |
| |
|
|
Table3: Anti-HIVactivityofpurinederivativesof4'-C-cyano-2'-deoxy-and4'-C-ethynyl-2'-deoxynucleosides. |
|
| |
| 1. Some of the purine derivatives of CNdN have high anti-HIV activity, but none of them gives an acceptable SI. |
| |
| 2. All the purine derivatives of EdN have both high anti-HIV activity and an acceptable SI. |
| |
| Anti-HIV activity of 4’SdN against drug-resistant HIV mutants |
| |
| Many 4’SdN showed very high anti-HIV activity against wild-type HIV. However, the most important point of our study is whether they are active against drug-resistant HIV-mutants. The anti-HIV activity of selected 4’SdN against HIV mutants resistant to various NRTI is listed in Table 4. |
| |
|
|
Table 4: Anti-HIVactivityofselected4'-C-substituted-2'-deoxynucleosideagainstwildtypeHIVanddrug-resistantHIV. |
|
| |
| It is noteworthy that the three cytidine derivtives maintained their activity against the drug-resistant HIV mutants, although the activity of 4’-C-ethynyl-D-arabinofuranosyl cytosine (EaraC) and MdC decreased significantly against M184V, M184I, and 41/69/125/SG. |
| |
| The three purine derivatives, 2’-deoxy-4’-C-ethynyladenosine (EdA), 2’-deoxy-4’-C-ethynyl-2-aminoadenosine (EAdA), and 2’-deoxy-4’-C- ethynylguanosine (EdG) except for 2’-deoxy-4’-C-ethynylinosine (EdI) were highly potent against all drug-resistant HIV mutants (EdI whih is not the component of DNA was much less active than the former three derivatives, especially against M184V). Additionally, the three were also active against a non-nucleoside reverse transcriptase inhibitor-resistant Y181C. Further, the three purine derivatives were highly potent against the HIV isolated from seven heavily drug-experienced patients with acquired immune deficiency syndrome (AIDS) as efficiently as against wild-type HIV [16,17,27]. Thus, EdA, EAdA, and EdG were highly potent against all the existing HIV. |
| |
| These results let us suppose that the three purine EdN derivatives could even prevent the emergence of drug-resistant HIV. It should be noted that EdG showed toxicity to Hela cells at 52μM, therefore, EdG would be highly toxic. |
| |
| Mouse toxicity of purine derivatives of EdN |
| |
| Since the three purine derivatives of EdN showed high activity against all HIV and acceptable SI, the mouse toxicity of these EdN was next examined [27]. |
| |
| All eight mice survived after a single dosage of 3~100mgkg-1 of EdA and EdI by both intravenous and oral administrations, but all mice died after a single dosage of 3mgkg-1 of EAdA and EdG irrespective of the administration method. Thus, it seemed that EdA and EdI were not toxic, but EAdA and EdG were highly toxic. |
| |
| However, in mice, it was found that EdA and EAA were easily converted to EdI and EdG, respectively, by adenosine deaminase [28]. The results showed that the actual toxicity of EdA and EAdA to animals is hard to estimate. |
| |
| Development of EdA derivatives stable to adenosine deaminase and their anti-HIV activity |
| |
| The fact that both EdA and EAdA are deaminated by adenosine deaminase prompted us to prepare EdA derivatives stable to the enzyme. It has been known that the adenine derivatives having a halogen atom at the 2-position of the base are stable to adenosine deaminase [28]. Therefore, at first, EFdA (Figure1) was synthesized and evaluated for anti-HIV activity [28-32] and stability to both adenosine deaminase and acidic conditions [30]. |
| |
| Because EFdA is a nucleoside derivative modified at two positions (4’-position and 2-position) of physiologic 2’-deoxyadenosine, the toxicity of EFdA is expected to be lower than that of EdA. As shown in Table 5, EFdA is highly potent against all HIV including multidrugresistant and M184V mutants and has an acceptable SI (110,000) [28,30-32]. |
| |
|
|
Table5: Anti-HIVactivityof2'-deoxy-4'-C-ethynyl-2-fluoroadenosine(EFdA). |
|
| |
| Expectedly, EFdA was completely stable to adenosine deaminase under the conditions where EdA was completely deaminated in 60min and, further, fairly stable under acidic conditions. Thus, in 120 min only a small part (3%) of EFdA was decomposed under the acidic conditions of gastric juices (pH 1.06) at 24°C, while 2’,3’-dideoxyadenosine (ddA) was completely decomposed in 5min [28,30]. |
| |
| Claims that 3'-OH of EdN is the cause of the toxicity of EdN |
| |
| While we were working on our project, two papers on the role of 3’- OH of 4’SdN appeared in which the authors claimed that 3’-OH was the cause of the toxicity of 4’SdN. Haraguchi and coworkers reported [33] that 4’-C-ethynyl-2’,3’-didehydro-3’-deoxythymidine (Ed4T) was more potent against wild-type HIV and multi-drug resistant HIV mutant but a little less potent against M184V-mutant and much less toxic than d4T. Further, they claimed that 3’-OH of 4’SdN was the cause of the toxicity. |
| |
| These results suggested that the ethynyl group of Ed4T has special affinity to RT and the additional modification of d4T by addition of ethyl group at the 4’-position decreased the toxicity of d4T. |
| |
| In contrast, Siddiqui and coworkers reported [34] that 3’-OH of 4’SdN played an important role in the phosphorylation of 5’-OH by cellular kinases, but was the cause of the toxicity of 4’SdN. This determination was based on their results that 4’-C-ethynyl-2’,3’- dideoxycytidine (EddC) was inactive against HIV in cellular systems, but its 5-O-triphosphate (EddCTP) was more active than 3’-azido-3’- deoxythymidine-5’-O-phosphate (AZTTP) against the RT of wild-type HIV. They also reported that EddCTP was much less active against RT of the M184V mutant than against the RT of wild-type HIV. In addition, the L-isomer of EddCTP was not active against the RT of the M184V mutant. |
| |
| Anti-HIV activity of 2’,3’-dideoxy (dd-) and 2’,3’-didehydrodideoxy (d4) analogs of EFdA |
| |
| The two papers previously cited caused us to synthesize the dd- and d4- analogs of EFdA and evaluate their anti-HIV activity [28,35]. |
| |
| The anti-HIV activities of 2’,3’-dideoxy-4’-C-ethynyl-2- fluoroadenosine (EFddA), 2’,3’-didehydrodideoxy-4’-C-ethynyl- 2-fluoroadenosine (EFd4A), and 2’-deoxy-4’-C-ethynyl-2- chloroadenosine (ECldA) together with that of Ed4T are listed in Table 6. Although both EFd4A and EFddA showed some activity against wild-type HIV, they significantly lost any activity against drug-resistant HIV. ECldA is highly active against all HIV, however, its activity is lower than that of EFdA. These results indicated that the 3’-OH played important roles not only for the phosphorylation of 5’-OH, but also for the activity against drug-resistant HIV. Further, these results together with the fact that Ed4T is more active than d4T, and further that 2’-deoxy-4’-C-ethyny-2- fluoroadenosine-5’-O-triphosphate (EFdATP) is two tomes better substrate than the physiologic 2’-deoxy adenosine- 5’-O-triphosphate (dATP) [36] indicated that the 4’-C-ethynyl group will have the special affinity to RT. Recently, it was reported by two groups [36,37] that the active center of the RT of HIV has a narrow hydrophobic cavity to accept preferably the ethynyl group, thus 4’-C-ethynyl nucleoside derivatives have highly potent anti-HIV activity. |
| |
|
|
Table 6: Anti HIV activity of 4’-C’- substituted-2’-deoxy-2-haloadenosines. |
|
| |
| Toxicity of EFdA to mice and inhibition of DNA polymerases |
| |
| Because EFdA is stable to adenosine deaminase, its mouse toxicity was examined [28-32]. EFdA did not show any acute toxicity to mice by either oral or intravenous administration up to 100mgkg-1. |
| |
| It is known that the toxicity of NRTI to animals is caused by their inhibition of mitochondrial DNA polymerase γ. The 50% effective concentration (EC50) of EFdATP to inhibit the incorporation of dATP mediated by human mitochondrion DNA polymerase was 10μM, which was significantly higher than the 0.2μM of 2’,3’-dideoxyadenosine-5-Otriphosphate (ddATP) [28,31] which is the real active form of ddI. The EC50 values of EFdATP against DNA polymerase α and β were higher than 200μM. These results indicate that the DNA polymerases scarcely recognize EFdATP, a derivative modified at two positions of physiologic dATP, as their substrate but that RT does. |
| |
| Intracellular metabolism of EFdA [31,35] |
| |
| The amounts of all fractions of intracellular EFdA metabolites, (EFdA-monophosphate, EFdA-diphosphate, and EFdATP) increased proportionately with an increase in the concentration of intracellular EFdA, while compared to AZT diphosphate and AZTTP, only AZT monophosphate markedly increased with an increase in intracellular AZT concentration. The intracellular half-life (T1/2) of EFdATP was ~18 h in complete expansion media (CEM) cells, MT4 cells, and multinuclear activation of galactosidase indicator (MAGI)-CCR5 cells (T1/2 of AZTTP was ~3 h). About 50% of the cells were protected against the infection of HIV for 24 h after removal of extracellular EFdA in both MT4 cells and MAGI cells cultured in the presence of 0.1μM of EFdA. |
| |
| These results indicate that EFdA, 2’-deoxy-4’-C-ethynyl-2- fluoroadenosine- 5-O-diphosphate (EFdADP), and EFdATP are very stable against intracellular enzymatic catabolism. |
| |
| Mechanism of inhibition of HIV-RT by EFdA |
| |
| The one site modified nucleosides, 4’-C-methyl-2’-deoxycytidine (MdC), 4’-C-ethynyl-2’-deoxycytidine (EdC), and 4’-C-ethynyl-2’- deoxyguanosine (EdG) are highly anti-HIV active and highly toxic, too. MdC was shown to be a chain-terminator of DNA polymerase [7]. These results showed that both DNA-dependent DNA polymerases and RT (RNA-dependent DNA polymerase) accept these one site modified nucleosides and the inhibitory activity could be ascribed to the unreactivity of the neopentyl type 3’-OH group. |
| |
| On the other hand, the mechanism of the supremely high anti- HIV activity of EFdA was rationalized by E. Michailidis and his coworkers as follows [37]. The special high affinity of EFdA to RT by both 4’-C-ethynyl group and 3’-OH makes the primer having a 3’-terminal EFdA-MP difficult to translocate from the pre-translocation site to the post-translocation site and therefore the next complementary nucleotide can’t react with the 3’-EFdA-MP. Thus, they named EFdA translocation-defective reverse transcriptase inhibitor (TDRTI). |
| |
| An improved synthesis of EFdA |
| |
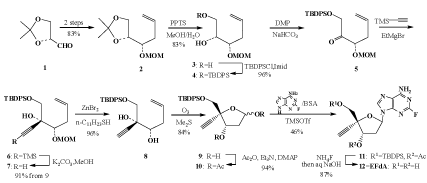
| The most critical point of our study on EFdA is the difficulty of the synthesis of EFdA. Our previous synthesis of EFdA took 18-steps with an overall yield of 2.5% starting from expensive 2-amino-2’- deoxyadenosine. Therefore, we developed a new 12-steps synthesis that gives EFdA in 18 % overall yield starting from much less expensive (R)- glyceraldehyde acetonide (Scheme 1) [38]. |
| |
|
|
Scheme 1: An improved synthesis of EFdA. |
|
| |
| Summary |
| |
| A supremely high anti-HIV active and lowly toxic EFdA which could prevent the emergence of resistant HIV mutants was developed by the studies based on the author’s working hypotheses. |
| |
| Acknowledgments |
| |
| The work described in this paper was achieved through the tremendous efforts of the many coworkers whose names appear in the cited references. |
| |
| The author wishes to express his sincere thanks to Prof. Dr. Hiroaki Mitsuya of Kumamoto University and NIH, Prof. Dr. Masanori Baba of Kagoshima University, Prof. Dr. Mineo Saneyoshi of Teikyo University for the biological studies of 4’SdN, and Prof. Dr. Shigefumi Kuwahara for the synthetic study of EFdA. This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan, Asahi Breweries LTD, and Yamasa Corporation. |
| |
| |
| References |
| |
- Mitsuya H, Weinhold K, Furman P, St Clair M, Lehrman S, et al. (1985) 3'-Azido-3'-deoxythymidine (BW A509U): an antiviral agent that inhibits the infectivity and cytopathic effect of human T-lymphotropic virus type III/lymphadenopathy-associated virus in vitro. Proc Natl Acad Sci U S A 82: 7096–7100.
- Murphy EL, Collier AC, Kalish LA, Mintz L, Wallach FR, et al. (2001) Highly Active Antiretroviral Therapy Decreases Mortality and Morbidity in Patients with Advanced HIV Disease. Ann Int Med 135: 17-26.
- Hung CC, Hsiao CF, Chen MY, Hsieh SM, Chang SY, et al. (2006) Improved Survival of Persons with Human Immunodeficiency Virus Type 1 Infection in the Era of Highly Active Antiretroviral Therapy in Taiwan. Jpn J Infect Dis 59: 222-228.
- Kirby KA, Singh K, Michailidis E, Marchand B, Kodama EN, et al. (2011) The Sugar Ring Conformation of 4’-Ethynyl-2-Fluoro-2’-Deoxyadenosine and Its Recognition by the Polymerase Active Site of HIV Reverse Transcriptase. Cell Mol Biol (Noisy-le-grand) 57: 40-46.
- Shuhadonlnik RJ (1970) Nucleoside Antibiotics, Wiley-International.
- Waga T, Ohrui H, Meguro H (1996) Synthesis and Biological evaluation of 4’-C-methyl nucleoside. Nucleosides and Nucleotides 15: 287-304.
- Waga T, Nishizaki T, Miyakawa I, Ohrui H, Meguro H (1993) Synthesis of 4'-C-methylnucleosides. Biosci Biotechnol Biochem 57: 1433-1438.
- Yamaguchi T, Tomikawa A, Hirai T, Kawaguchi T, Ohrui H, et al. (1997) Antileukemic Activities and Mechanism of Action of 2’-Deoxy-4’-Methylcytidine and Related Nucleosides. Nucleosides and Nucleotides 16: 1347-1350.
- Waga T, Nishizaki T, Miyakawa I, Ohrui H, Meguro H (1993) Synthesis of 4’-C-methyl nucleosides. Biosci Biotechnol Biochem 57: 1433-1438.
- Youssefyeh RD, Verheyden JP, Moffatt JG (1979) Synthesis of 4’-C-hydroxymethyl ribofuranosyl nucleosides. J Org Chem 44: 1301-1321.
- Kitano K, Miura S, Ohrui H, Meguro H (1997) Synthesis of 4’-C-Fluoromethylnucleosides as Potential Antineoplastic Agents. Tetrahedron 53: 13315-13322.
- Kitano K, Machida H, Miura SS, Ohrui H (1999) Synthesis of Novel 4’-C-Methyl-Pyrimidine Nucleosides and Their Biological Activities. Bioorg Med Chem Lett 9: 827-830.
- Kohgo S, Horie H, Ohrui H (1999) Synthesis of 4’-C-Ethynyl-ß-D-arabino- and 4’-C-Ethynyl-2’-deoxy-ß-D-ribo- pentofuranosyl Pyrimidines, and their biological evaluation. Biosci Biotechnol Biochem 63: 1146-1149.
- Kohgo S, Kodama E, Shigeta S, Saneyoshi M, Machida H, et al. (1999) Synthesis of 4’-substituted nucleosides and their biological evaluation. Nucleic Acids Symp Ser 42: 127-128.
- Ohrui H, Kohgo S, Kitano K, Sakata S, Kodama E, et al. (2000) Synthesis of 4’-C-Ethynyl-ß-D-arabino- and 4’-C-Ethynyl-2’-deoxy-ß-D-ribo- pentofuranosyl pyrimidines and –purines and Evaluation of their anti-HIV activity. J Med Chem 44: 4516-4625.
- Kohgo S, Mitsuya H, Ohrui H (2001) Synthesis of the L-Enantiomer of 4’-C-Ethynyl-2’-deoxycytidine. Biosci Biotechnol Biochem 65. 1879-1882.
- Kodama E, Kohgo S, Kitano K, Machida H, Gatanaga H, et al. (2001) 4’-Ethynyl Nucleoside Analogs: Potent Inhibitor of Multidrug-Resiatant Human Immunodeficiency Virus Variants In Vitro. Antimicrob Agents Chemother 45: 1539-1546.
- Ohrui H, Mitsuya H (2001) 4’-C-Substituted-2’-Deoxynucleosides: A Family of Antiretroviral Agents Which are Potent against Drug-Resistant HIV Variants. Curr Drug Targets Infect Disord 1: 1-10.
- Maag H, Rydzewski RM, McRoberts MJ, Crawford-Ruth D, Verheyden JP, et al. (1992) Synthesis and anti-HIV activity of 4’-azido- and 4’-methoxynucleosides. J Med Chem 35: 1440-1451.
- O-Yang C, Wu HY, Fraser-Smith EB, Walker KAM (1992) Synthesis of 4’-cyanothymidine and analogues as potent inhibitor of HIV. Tetrahedron Lett 33: 37-40.
- O-Yang C, Kurz W, Eugui EM, McRoberts MJ, Verheyden JPH, et al. (1992) 4’-Substitited nucleosides as inhibitors of HIV: an unusual oxetane derivative. Tetrahedron Lett 33: 41-43.
- Prisbe FJ, Verheyden JPH, Rydzewski RM (1993) Nucleosides & Nucleotides as Antitumor & Antiviral Agents. Edited by Chu C. K. & Baker. D. C. New York: Plenum Press, pp. 101.
- Chen MS, Suttmann RT, Papp E, Cannon PD, McRoberts MJ, et al. (1993) Selective action of 4’-azidothymidine triphosphate on reverse transcriptase of human immunodeficiency virus type 1 and human DNA polymerase alpha and beta. Biochemistry 32: 6002-6010.
- Wang G, Seifert WE (1996) Synthesis and evaluation of oligodeoxynucleotides containing 4'-C-substituted thymidines. Tetrahedron Lett 37: 6515-6518.
- Nomura M, Shuto S, Tanaka M, Sasaki T, Shigeta S, et al. (1999) Nucleosides and nucleotides. 185. Synthesis and biological activities of 4'-alpha-C-branched-chain sugar pyrimidine nucleosides. J Med Chem 42: 2901-2908.
- Beach JW, Jeong LS, Alves AJ, Pohl D, Kim HO, et al. (1992) Synthesis of enantiomerically pure (2'R,5'S)-(-)-1-(2-hydroxymethyloxathiolan-5-yl) cytosine as a potent antiviral agent against hepatitis B virus (HBV) and human immunodeficiency virus (HIV). J Org Chem 57: 2217-2219.
- Kohgo S, Yamada K, Kitano K, Iwai Y, Sakata S, et al. (2004) Design, Efficient Synthesis, and Anti-HIV Activity of 4'-C-Cyano- and 4'-C-Ethynyl-2'- deoxy Purine Nucleosides. Nucleosides Nucleotides Nucleic Acids 23: 671-690.
- Hayakawa H, Kohgo S, Kitano K, Ashida N, Kodama E, et al. (2004) Potential of 4'-C-substituted nucleosides for the treatment of HIV. Antivir Chem Chemother 15: 169-187.
- Patent: International Publication Number, 2005/090349, PCT/JP2005/005374.
- Montgomery JA, Hewson K (1969) Nucleosides of 2-fluoroadenine. J Med Chem 12: 498-504.
- Ohrui H, Kohgo S, Kitano K, Hayakawa H, Kodama E, et al. (2005) 2’-deoxy-2-fluoroadenosine, a nucleoside derivative potent against HIV-1 with no acute mouse toxicity,: Highlights of the role of 3’-OH for biological activity. 4’-C-Ethynyl-Abstract of the 18th International Conference on Antiviral Research. Barcelona April pp.16.
- Nakata H, Amano M, Koh Y, Kodama E, Yang G, et al. (2007) Activity against human immunodeficiency virus type 1, intracellular metabolism, and effects on human DNA polymerases of 4'-ethynyl-2-fluoro-2'-deoxyadenosine. Antimicrob Agents Chemother 51: 2701-2708.
- Kawamoto A, Kodama E, Sarafianos SG, Sakagami Y, Kohgo S, et al. (2008) 2'-Deoxy-4'-C-ethynyl-2-halo- adenosines active against drug-resistant human immunodeficiency virus type 1 variants. Int J Biochem Cell Biol 40: 2410-2420.
- Haraguchi K, Takeda S, Tanaka H, Nitanda T, Baba M, et al. (2003) Synthesis of a highly active new anti-HIV agent 2', 3'- didehydro-3'-deoxy-4'-ethynylthymidine. Bioorg Med Chem Lett 13: 3775-3777.
- Siddiqui MA, Hughes SH, Boyer PL, Mitsuya H, Van QN, et al. (2004) 4'-C-Ethynyl-2',3'-dideoxynucleoside Analogue, Highlights the Role of the 3'-OH in Anti-HIV Active 4'-C-Ethynyl-2'-deoxy Nucleosides. J Med Chem 47: 5041-5048.
- (a) Ohrui H (2011) Development of modified nucleosides that have supremely high anti-HIV activity and low toxicity and prevent the emergence of resistant HIV mutants. Proc Jpn Acad Ser B Phys Biol Sci 87: 53- 65.
(b) Ohrui H (2006) 2'-deoxy-4'-C-ethynyl-2-fluoroadenosine, a nucleoside reverse transcriptase inhibitor, is highly potent against all human immunodeficiency viruses type 1 and has low toxicity. Chem Rec 6: 133-143.
- Young G, Wang J, Cheng Y, Dutschmam GE, Tanaka H, et al. (2008) Mechanism of Inhibition of Human Immunodeficiency Virus Type 1 Reverse Transcriptase by a Stavudine Analog, 4’-Ethynyl Stavudine Triphospaphate. Antimicrob Agents Chemother 52: 2035-2042.
- Michailidis E, Marchand B, Kodama EN, Singh K, Matsuda M, et al. (2009) Mechanism of Inhibition of HIV-1 Reverse Transcriptase by 4'-Ethynyl-2-fluoro-2'-deoxyadenosine Triphosphate, a Translation -defective Reverse Transcriptase Inhibitor. J Biol Chem 18: 35681-35691.
- Kageyama M, Nagasawa T, Yoshida M, Ohrui H, Kuwahara S (2011) Enantioselective Total Synthesis of Potent Anti-HIV Nucleoside EFdA. Org Lett 13: 5264-5266.
- Eldrup AB, Prhavc M, Brooks J, Bhat B, Prakash TP, et al. (2004) Structure-activity relationship of heterobase-modified 2'-C-methyl ribonucleosides as inhibitors of hepatitis C virus RNA replication. J Med Chem 47: 5284-5297.
- Klumpp K, Kalayanov G, Ma H, Pogam SL, Leveque V, et al. (2007) 2’-Deoxy-4’-azido Nucleoside Analogs Are Highly Potent Inhibitors of Hepatitis C Virus Replication Despite the Lack of 2’-a-Hydroxyl Groups. J Biol Chem 14: 2167-2175.
- Smith DB, Kalayanov G, Sund C, Winqvist A, Pinho P, et al. (2009) The Design, Synthesis, and Antiviral Activity of 4#-Azidocytidine Analogues against Hepatitis C Virus Replication: The Discovery of 4’-Azidoarabinocytidine. J Med Chem 52: 219-223.
- Hattori H, Tanaka M, Fukushima M, Sasaki T, Matsuda A (1996) Nucleosides and Nucleotides. 158. 1-(3-C-Ethynyl-ß-D-ribo-pentofuranosyl)-cytosine, 1-(3-C-Ethynyl-ß-D-ribo-pentofuranosyl)uracil, and Their Nucleobase Analogs as New Potential Multifunctional Antitumor Nucleosides with a Broad Spectrum of Activity. J Med Chem 39: 5005-5011.
- Ohrui H (2009) A proposed structure of modified nucleosides expected to have high antiviral activity and low toxicity. Nucleosides Nucleotides Nucleic Acids 28: 772-775.
|
| |
| |