| Research Article |
Open Access |
|
| Marin I Danciu* |
| Street PetreBanita, Number 204, Calarasi Postcode 207170, Judetul Dolj, Country Romania |
| *Corresponding authors: |
Marin I Danciu
Street Petre Banita
Number 204, Calarasi Postcode 207170
Judetul Dolj, Country Romania
Tel: +40721183450
E-mail: danciu@prodanciu.ro |
|
| Â |
| Received December 09, 2011; Published October 25, 2012 |
| Â |
| Citation: Danciu MI (2012) Danciu Laws of Heredity-Four New Laws. 1:447. doi:10.4172/scientificreports.447 |
| Â |
| Copyright: © 2012 Danciu MI. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. |
| Â |
| Abstract |
| Â |
| By Mendel laws the way of transmitting characters from parents to descendants was less clarified, and in some regards (dominance, recessiveness, sex chromosomes, gametes endowment, transmitting some characters to the 1st, the 2nd, the 3rd generation etc.) consistent errors were made. Also, a good inspiration led to the creation (Figure 1) and once made this graph, was easy to see that it suggests the existence of several rules by which the characters from parents to their descendants are transmitted. So, based on this chart, are discovered and formulated four new laws of heredity: the law of gametes’ contribution, the law of sex chromosomes, the law of hereditary autosomal factors, and the law of gametes endowment. |
| Â |
| Danciu laws of heredity: Eliminate all errors of heredity made so far; completily clarifies the way characters are transmitted from parents to descendants, including most the autosomal - recessive diseases; are valid 100% in all cases and all species that reproduce by gametes; can have a significant practical importance in the creating activity of plants and animals hybrids with a high biological value; so, by a simple correlation of the moment of crosses/ pairing with the appearance of the mature gametes, hybrids can be obtained with the most active hereditary factors from the most valuable individual; can be an important tool in the future research in genetics and mostly in the medical genetic research. |
| Â |
| Keywords |
| Â |
| Autosomal; Characters transmitted; Crossing-Over; Gametes; Recessive; Sex chromosomes; Laws of heredity |
| Â |
| Introduction |
| Â |
| By Mendel laws the way of transmitting characters from parents to descendants was less clarified, and in some regards (dominance, recessiveness, sex chromosomes, gametes endowment, transmitting some characters to the 1st, 2nd, the 3rd generation etc.) consistent errors were made. Also, for to find out why most girls resemble more with their fathers and most boys resemble more with their mothers a good inspiration led to the creation (Figure 1). Once made this chart, was easy to see that it suggests the existence of several rules by which the characters from parents to their descendants are transmittedtransmitted. Based on this chart, are discovered and formulated four new laws of heredity: the law of gametes’ contribution, the law of sex chromosomes, the law of hereditary autosomal factors, and the law of gametes endowment. |
| Â |
|
|
| Â |
| Methods |
| Â |
| Practical observations of reality (to humans, plants, animals); Multidisciplinary theoretical studies. |
| Â |
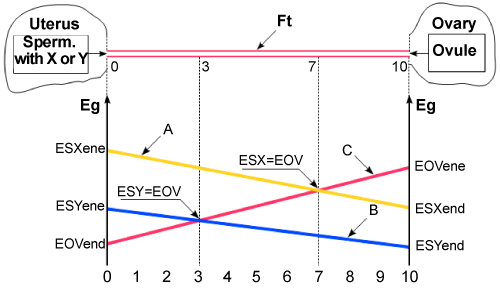
| Danciu chart (Figure 1) |
| Â |
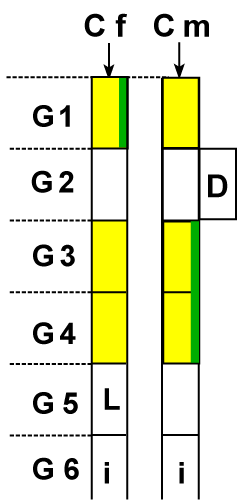
| Explanations: (Figure 1) At the moment of the entrance in the fallopian tube, the sperm (with X or Y chromosome) and the ovule have an energy of a certain size, noted on the chart with ESXene, ESYene and EOVene. This energy, they had at the entrance, is decreasing in time. The decrease of the energy is represented by A and B lines, for the two sperm and by C line for the ovule. At the same time, the gametes occupy new positions in the tube, each of them coming in contact with the other one and then them fusion. The contact between the sperm and the ovule (fecundation) can take place in any point on the entire length of the tube. At the meeting point, the energies of the two gametes are different in size, so there is a difference between them. This energy difference, in the favour of one gamete or of the other, exists in all the points on the length of the tube, except for one point. This is the point when the energies of the two gametes are equal. Before this point, the difference of energy is in favour of one gamete and after this point the difference is in favour of the other gamete. (Point 7 in the figure 1 where the lines A and C join and point 3, where lines B and C join). Following the chart, it can be easily understood, when and how much the child will resemble more one of the parents (or/and one of his ancestors). |
| Â |
| The case of boy-child, when in fecundation the spermatozoon that contains Y chromosome participates |
| Â |
| If the fecundation takes place on the 0-3 segment of the tube, where the energy ESY of the sperm is bigger than the energy EOV of the ovule, the boy will resemble more the father, because the energy difference is in favour of the sperm. The resemblance is more emphatic when the fecundation takes place closer to the uterus (towards 0 point), where the energy difference is bigger. If fecundation takes place on the segment 3-10 of the tube, where the energy EOV of the ovule is bigger than the energy ESY of the sperm., the boy will resemble more the mother, because the energy difference is in favour of the ovule. The resemblance is more emphatic when the fecundation takes place closer to the ovary (towards point 10), where the energy difference is bigger). |
| Â |
| If fecundation takes place in point 3, then the resemblance is 50% the father and 50% the mother, because all the energies are equal, ESY= EOV. |
| Â |
| Do observe that segment 3-10 of the tube is longer (almost 70% of the total) than the segment 0-3 of the tube (almost 30% of the entire length). This means that the probability of the fecundation to take place on 3-10 segment of the tube is bigger than on segment 0-3. So, it is more likely that boys should resemble their mothers more than their fathers. This fact is in accordance with reality, where anyone can notice that most boys (almost 70% of their number) resemble more their mothers and fewer boys (almost 30%) resemble their fathers. |
| Â |
| The case of the girl-child, when in fecundation participates X type spermatozoon |
| Â |
| If fecundation produces on the 0-7 segment of the tube, where the energy ESX of the sperm is bigger than the energy EOV of the ovule, the girl will resemble more her father. This will happen because the energy difference is in favour of the spermatozoon. The resemblance is more emphatic as the fecundation produces closer to the uterus (towards point 0), where the energy difference is bigger. |
| Â |
| If fecundation produces on the segment 7-10 of the tube, where the energy EOV of the ovule is bigger than the energy ESX of the sperm, the girl will resemble more the mother, because here the energy difference is in favour of the ovule. The resemblance is much more emphatic as the fecundation produces closer to the ovary (towards point 10), where the energy difference is bigger. |
| Â |
| If fecundation takes place in point 7, then the resemblance is 50% the father and 50% the mother, because here the energies are equal, ESX=EOV. |
| Â |
| Do observe that segment 0-7 of the tube is longer than the segment 7-10 (approximate 70% and 30% of the length of the tube). It means the probability that fecundation produces on the 0-7 segment of the tube is bigger than on 7-10 segment. So, the probability that girls resemble the fathers more is bigger than the probability that they resemble their mothers more. This fact is in accordance with reality where anyone can notice that most girls (almost 70% of their number) resemble more their fathers and fewer girls (almost 30%) resemble their mothers. |
| Â |
| With the explanations above, was demonstrated that the (Figure 1) entirely reflects reality. |
| Â |
| From the (Figure 1) we could grasp three extremely important conclusions: any gamete contains the entire set of hereditary factors (the entire information) necessary for the complete edification of a new organism, in regular health conditions; the participation, with hereditary factors, of each of the two gametes to the complete edification of the new organism is complementary, meaning the sum of their contributions, expressed in %, is equal with 100%; in principle, the (Figure 1) is valid for all species that reproduce by gametes. |
| Â |
| Results and Discussion |
| Â |
| Law 1: The law of gametes' contribution |
| Â |
| The contribution with hereditary factors of the gametes, to the content of the set of n active chromosomes, is determined by the energy that the same gametes have at the moment of fecundation. The size of the contribution with hereditary factors of one gamete depends on the energy of both gametes. For each gamete, the size of the contribution with hereditary factors that will be admitted in the set of n active chromosomes, will be in such a way that the proportion of the contribution in the set be equal to the proportion of the energy of respectively gamete in the sum of the energies of both gametes. For each gamete, the proportion of the contribution with factors that will be admitted in the set of n active chromosomes is also the proportion of the contribution in the set with factors that will be effectively activated (that will determine the phenotype, between all the admitted ones). Hereditary factor, expression introduced by Mendel, defines any carrier of hereditary information: whole chromosome (this is the highest rank factor); segment of chromosome (with two or more genes); whole gene; segment of gene (with two or more nucleotides); nucleotide (this is the lowest rank factor). "n" represents the total number of chromosomes of a gamete. For the full understanding of this law, beside the (Figure 1), the following example (hypothetical) is probatory. At the moment of contact for fecundation, the sperm has 120 energy units and the ovule 80, the entire assembly has 200 energy units. In the amount of 200, 120 represents 60% and 80 represent 40%. So, 60% of the hereditary factors of the set of n active chromosomes represent the sperm’s contribution and 40% the ovule’s contribution. The child will resemble the father more, because the highest quantity of hereditary factors comes from the spermatozoon. The (nascent) egg cell does not operate with numbers, as in this example, but it has proteins-enzymes systems that recognize the gametes, evaluate their energies and configure the set of n active chromosomes, following precisely the proportions issued out according to the law 1. |
| Â |
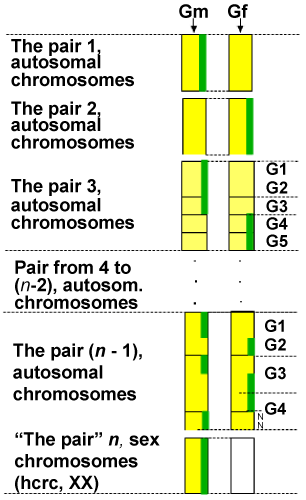
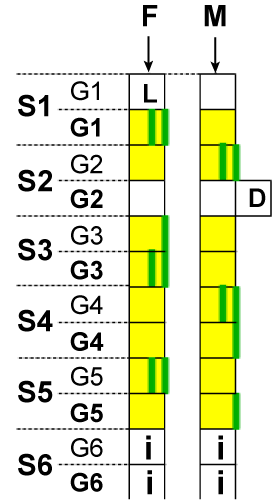
| According to law 1, just the quantitative contribution of each gamete is established and not the factors that effectively make this contribution. From this point of view, the conclusion is that the configuration process of the set of n active chromosomes, by the proteins-enzymes, takes place according to the fundamental principle specific to all natural systems, so to the biological ones also, named the principle of the highest efficiency, meaning, the lowest energy consumption. According to this principle, the configuration (by marking factors) of the set is always made by choosing that option which requires the lowest energy consumption, between the many available options. The homologues of the marked factors, so the homologues of the ones admitted in the set of n active chromosomes, form, as a consequence, the set of n inactive chromosomes. Once configured, the sets maintain the same structure in all somatic cells, including in the gametes forming cells, during the entire life time of that respectively organism. By marking, the hereditary factors admitted in the set of n active chromosomes are not dislocated from the positions they occupy in their original gametes. The effective configuration process, and thus of marking the chosen factors to be part of the n active chromosomes set, starts mandatory with the sex chromosome, X or Y, always both of them from the male gamete only (according to law 2, for X and Y, and 3, for autosomes). The sex chromosome is first admitted for the following reasons: the set of n active chromosomes cannot be a complete set, unless it has a sex chromosome; the size of the contribution of each gamete, resulted at the evaluation, is possible to be strictly respected only by fragmentation of some factors, or the sex chromosome cannot be fragmented, because between X and Y there is no homology to permit the mutual completion without affecting the purity of sex. Therefore, the total elimination of the fragmentation risk is possible only if the sex chromosome is admitted first and mandatory whole, to exclude the sex mixture. From both gametes, other whole chromosomes, segments of chromosomes, whole genes follow up, and segments of genes are admitted (marked) for the precise placement in the contribution proportion resulted at the evaluation, and for the smooth tuning, even nucleotides are admitted. This is the mechanism by resulting two sets of n, active and inactive, chromosomes, both containing whole chromosomes (physically) as well as combined chromosomes, each combined chromosome having factors (low or high rank) physically situated on both pair-chromosomes. The above statements can be easily grasped from (Figure 2). In this example, the set of n active chromosomes is made up of the factors marked with a green line. So, we have: X sex chromosome from the of male gamete (from n “pairâ€); whole chromosome from the pair 1, of male gamete; whole chromosome from the pair 2, of female gamete; a segment of chromosome made up of 3 genes (G1, G2, G3) from the pair 3, of the male gamete; a segment of chromosome made up of 2 genes (G4, G5) from the pair 3, of the female gamete; other factors from the male and female gamete in pairs 4 to (n-2) and which do not appear in the chart; a whole gene (G1) from pair (n-1), from the male gamete; a whole gene (G2) from pair (n-1), from the female gamete; a segment of the gene (G3) from pair (n-1), from the male gamete; a segment of the gene (G3) from pair (n-1), from the female gamete; a segment of the gene (G4) from pair (n-1), from the female gamete; two nucleotides (N1 and N2) of the gene G4 from pair (n-1), from the male gamete; In this chart, the active chromosome in pair 3 and the active chromosome in pair (n-1) are combined chromosomes, in functional aspect, having in their structure factors, physically situated on both pair-chromosomes. |
| |
| Â |
|
|
Figure 2: Example of configurating the set of n active chromosomes |
|
| Â |
| Based on the law of gametes' contribution other conclusions can be drawn. We now know very clear what exactly determines one character to be dominant. This way, to any species and individual, any character can be dominant, if the hereditary encoder factor is chosen to be part of the set of n active chromosomes, based on the energy of the two gametes, from the moment of fecundation, and of the principle the lowest energy consumption, and (law 3) if the respectively factor is chosen to be effectively activated, so they will express. Based on this law, the concept called recessive, no longer refers to any factor that it is not expressed; the homologues of the autosomal factors admitted in the set of (n-1) active autosomal chromosomes cannot be called recessive any longer, because they were already scheduled not to express themselves. Would be more appropriate to be called surplus factors, because they are not necessary for the complete edification of the organism. A character can be called recessive only if the encoding factor cannot express, although it should, the result of this non-functioning is what was called the autosomal-recessive disease. The law of gametes' contribution can have a significant practical importance in the creating activity of plants and animals hybrids with a high biological value. By a simple correlation of the moment of crosses/pairing with the appearance of the mature gametes, hybrids can be obtained with the most active hereditary factors from the most valuable individual. The great advantage for mankind would be getting the food that to be, maybe not the greatest quantities, but certainly healthy [1-11]. |
| Â |
| Law 2: The law of sex chromosomes |
| Â |
| Sex chromosomes from the male body: The type Y chromosome originates from the father and it is a component of the set of n active chromosomes in the somatic cell of the male, having the role of determining the specific characters for the masculine gender. It is transmitted by one of the male-brothers gametes from one generation to another only to male descendant and always having the capacity of being active in their bodies. |
| Â |
| The type X chromosome originates from the mother and is part of the set of n inactive chromosomes in the somatic cell of the male. It is transmitted by the other male-brother gamete from one generation to another only to female descendants and always having the capacity of being active in their bodies. |
| Â |
| Sex chromosomes in female body: The type X chromosome originating from the father in the somatic cell of the female it is part of the set of n active chromosomes, having the role of determining the specific characters to feminine gender. It is transmitted by a female-brother gamete, from one generation to the other to male and female descendants and always lacking the capacity of being active in their bodies. |
| Â |
| The type X chromosome originating from the mother in the somatic cell of the female, it is part of the set of n inactive chromosomes. It is transmitted by the other female-brother gamete from one generation to the other to male and female descendants and always lacking the capacity of being active in their bodies. |
| Â |
| Each of the above mentioned four sex chromosomes (Y, X, X, X) carries its own and specific hereditary information, originating from ancestors from the origin of lines: one male, for Y, and three females, one for each X. This information, in regular health conditions, is transmitted unmodified from one generation to the other and is different from the one of similar chromosomes originating from other transmitters (genitors), from the origin of other lines. In the process of edification for a new organism, among all the existing hereditary factors on the sex chromosome, normally, only one of the factors (copies) that have the same functional role will be effectively activated so it will be expressed. |
| Â |
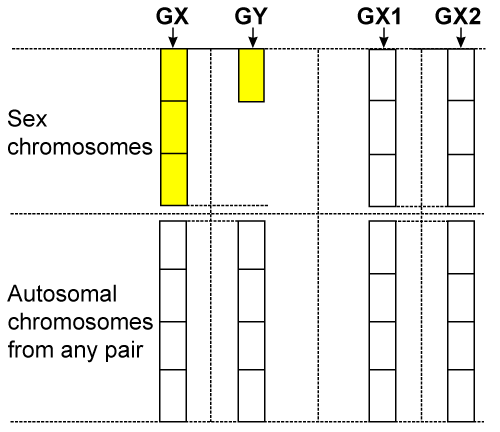
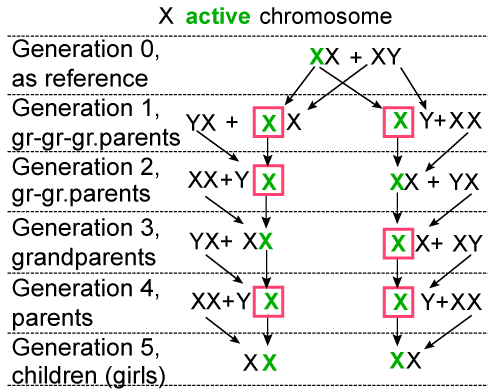
| The essence of this law consists in only sex chromosomes transmitted by the father are active in the children’s body: Y in the body of boys and X in the body of girls; the two X chromosomes transmitted by the mother are always lacking the capacity of being active, both in the body of boys and girls. The validity of this law results from the following reasoning. In order to become functional, any chromosome takes orders only from the protein-enzymes systems. These systems control all the processes, including the activation of chromosomes. But their orders can only be received by the (factors) chromosomes that already have the capacity to be active, and during the gamete phase only the female gamete contains protein-enzymes systems. As a result, for the safety of reproduction, protein-enzymes systems cannot exist in the same location (female gamete) with chromosomes (factors) that already have the capacity to be active. Thus, the possibility of (accidental) activation of chromosomes in the phase of female gamete was excluded, fact that reflects reality: no cases of female self-reproduction, from unfertilized ovules, are known. This law invalidates the concept regarding the inactivation of one of the two X chromosomes of the somatic cell of the female. At the moment of fecundation, there are no X chromosomes already active, in order for one of them to need to be inactivated; there is only one X chromosome having the capacity to be active, provided by the father and one without having the capacity to be active, provided by the mother. Therefore, between XY chromosomes at the male and XX at the female there is no scheduled functional interdependence nor can there be one; each of these chromosomes contains the entire information in order to accomplish, single, its role. For a better understanding of the law see as relevant (Figures 3 and 4) [12-24]. |
| Â |
|
|
Figure 3: The state of the sex chromosomes in the gametes |
|
| Â |
|
|
Figure 4: Example of transmitting the X active chromosome |
|
| Â |
| Law 3: The law of hereditary autosomal factors: Up until the moment of fecundation, the hereditary autosomal factors of the gametes (male and female) do not have the capacity of being active in a new organism; the exception are the autosomal factors attached by accident to the X chromosome transmitted by the father, these factors being already capacitated during the meiosis, together with the X chromosome. Normally, the capacity of being active is acquired (by mutual capacitation) during the fecundation, but only by those hereditary autosomal factors, which, after the fusion of the gametes (male and female), fulfil two conditions; first: they are present, at the level of the same position (locus), only as a unique pair of two homologue factors, one from the male gamete and one from the female gamete; second: the two homologue factors of any pair, should not be identical (not contain the same hereditary information), so, it should not originate from the same genitor (common ancestor). |
| Â |
| Of all the hereditary autosomal factors that acquired the capacity to be active (in the conditions mentioned) are admitted in the set of (n-1) active autosomal chromosomes, only one from each pair. Their homologues compose the set of (n-1) inactive autosomal chromosomes. |
| Â |
| In regular conditions, in the process of edification of a new organism, of all the admitted factors in the set of (n-1) active autosomal chromosomes, only one of each factor (variants) that have the same functional role will be effectively activated, so they will express. |
| Â |
| The admittance of the hereditary factors, in the set of n active chromosomes and the establishment of the ones that will be effectively activated are made by the protein-enzymes that have these kinds of roles. These only come from the female gamete and take action in accordance with the law of gametes' contribution, the law of sex chromosomes and with the principle of the lowest energy consumption."1" represents the sex chromosome of the gamete. Normally, any gene has more variants and/or copies on the same chromosome, one on locus; each encodes completely the same character. The variants, coming from more genitors from the origin of lines, have different hereditary information and exist only on autosomal chromosomes. The copies coming from only one genitor are identical, exist on sex chromosomes and, rarely, on autosomal chromosomes. Probably, the maximum possible number of variants or copies of a gene is characteristic to a species, maybe even rase (variety). On the same chromosome, there are more factors (genes) with the same functional role, for the safety of reproduction, the reproduction being, seems, the supreme purpose of every living creature. The chances to reproduce are much bigger as every factor, still has enough backup variants (copies). The autosomal hereditary factors make the two sets of autosomal chromosomes, active and inactive, each set having (n-1) autosomal chromosomes. According to first condition (law 3), the presence at fecundation, at the level of the same position (locus), of homologue factors in a smaller or bigger than 2 number (valid pair), excludes the possibility that in the future organism any of those factors to be functional (to manifest). So, this is recessive. According to second condition, the presence of the identical factors, so of the factors that in ascendance belonged to the same common ancestor (parents, grandparents, grand-grandparents etc), also excludes the possibility that any of the factors to be functional in the future organism. This is also recessive. When the set of (n-1) active autosomal chromosomes contains more autosomal factors, located on different lochs, but which encode same traits, only one factor from all these factors that have a functional role will be activated (expressed). The other factors remain latent, although they are in the set of active chromosomes. But any of these can be chosen to be activated in a future generation, closer or farther, if it will be part of the set of active chromosomes in all generations up until that moment. This is how it is explained the resemblance of the child, in some ways, not with the parents, but with the grandparents or with the grand-grandparents or even with farther ancestors. If these two conditions mentioned in the law are not fulfilled, the result will be the non-functioning of that factor (from the simple nucleotide up to the entire chromosome), because they did not encounter the corresponding homologue, in order to realize a valid pair, in any position (locus) of that factor. Or, the non-functioning of that factor has as a consequence the lacking of the encoded character from the new organism. This is how it is explained the apparition of novo and/or the transmitting from parents to children of the autosomal-recessive diseases like hemophilia, Dalton disease and many others. For understanding correctly of this law, a few charts and supplementary explanations are necessary. Thus, if a random pair of autosomal chromosomes is shown the way they gain the capacity to be active and the way the set of n active chromosomes is made (Figure 5). |
| Â |
|
|
| Â |
| The identical G6 genes does not capacitate because: the effective consanguinity (consanguinity = 100%) was reached and it was proved that the living systems do not accept it; in fact, between the two factors taken as chemical species, there are and action electro-magnetic forces of rejection, forces that appear between identical molecular structures; there are the same forces that marked the separation of factors during mitosis. By following the chart and the text of law it results: G2 gene, being present in 3 items instead of 2 in order to form a pair, does not capacitate. As a consequence, it will not be admitted in the set of active chromosomes, so it will not be functional (it will be recessive); G5 gene on the chromosome from the father (does not contain) is lacking hereditary information; it only has a support, so the gene suffered a deletion. In this situation, G5 gene from the mother, although in regular parameters, does not have anything to form a valid pair with, so, it has nothing to capacitate. As a result, the G5 gene will not function either (it will recessive). G6 gene being identical (come from same genitor), will not function, according to what was mentioned above (will be recessive). From the valid, capacitated pairs, in the set of active chromosomes, only the genes marked with a green line, so only one from each pair are admitted (according to law 1 and to the principle of the lowest energy consumption). In the figure 6, it is shown how the action law in the case when the factors with same functional role are 2 on each segment of autosomal chromosome from random pair, so: at the level of gene G1 a valid pair is not made, as the gene from the father is missing; as a result, the gene G1 from the mother does not capacitate so, it will not be functional; the gene G2: because 3 genes present at the level of the same locus and not only 2 it will not be capacitated so, it will not be functional; the pairs G6 and G6 each represented by 2 identical genes (effective consanguinity) cannot have any functional gene, as none of them capacitates; all the other factors mutual capacitate at the level of same pair, then are admitted in the set of active chromosomes the ones marked with 1 green line (according to the laws 1 and 3, and to the principle the lowest energy consumption) among which, just the ones marked with the second green line will be effectively activated at the scheduled moment; so, it will be effectively activated just 1 factor at a time among the 2 with same functional role; we notice that the disease caused by the idleness of G1 factor does not happen, as the same role is played by the G1 back up from the father; at the same time, the disease caused by the idleness of G2 factor does not happen, same role being played by G2 from the mother; instead, it will manifest the disease caused by factor G6 (G6), because of the idleness of this factor. So, the combined chromosome (from functional point of view), from the set of active chromosomes formed right after fecundation according to laws 1 and 3 and to the lowest energy consumption principle, contains the following factors that will express (they will be effectively activated): G1, G3 and G5 genes on the chromosome from the father; G2 and G4 genes on the chromosome from the mother. Conclusion: the autosomal-recessive diseases only happen when factors with the same functional role cannot make together with their homologues, not even one valid pair, knowing that it should be enough a valid pair in order for the disease not to act. [8,25-28]. |
| Â |
|
|
| Â |
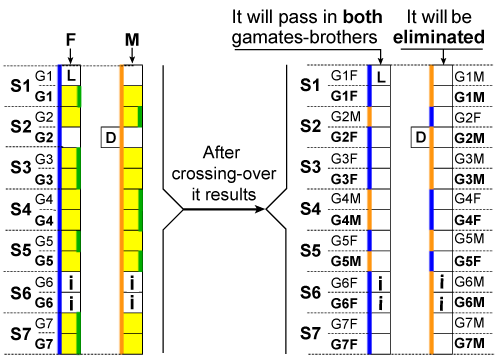
| Law 4: The law of gametes endowment: When forming the gametes (in meiosis), after separating by crossing-over the combined pair-chromosomes and doubling by mitosis the set of (n-1) autosomal active chromosomes, always both brothers-gametes receive, except for the sex chromosome, which is different, the same hereditary information, as follows: the set of n active chromosomes of that organism is entirely transmitted to one of the brothers-gametes; from the set of n inactive chromosomes, only the sex chromosome is transmitted to the other gamete, and, for completing the suite, a set of autosomal chromosomes identical with the ones of the gamete-brother is transmitted. |
| Â |
| If the set of (n-1) active autosomal chromosomes is incomplete, because of not admittance during the fecundation of some factors, they will be admitted now, during meiosis, in the set of the forming gametes and of the missing factors which can be (depending on the real cause of not admitting them and of the carrying chromosome which should be the next one to be transmitted): factors lacking hereditary information (they only contain supports), so abnormal factors, by deletion; normal factors; factors that contain two dozes of information on the same locus, so abnormal factors, by duplicating. |
| Â |
| The possible active hereditary factors accidentally attached to the inactive hereditary factors (during current meiosis) are not transmitted to the gametes. To any female gamete, also the protein-enzyme systems is transmited. They are designed to initiate and control all biochemical processes of after fecundation. |
| Â |
| Autosomal chromosomes of the set of (n-1) inactive autosomal chromosomes, being Surplus hereditary factors, are not transmitted to the gametes and they are eliminated; the eliminated chromosomes contain the eventual homologue factors of the ones that completed (during meiosis) the set of (n-1) autosomal chromosomes transmitted to the gametes. The possible inactive autosomal hereditary factors that accidentally attached, during current meiosis, to any of the n chromosomes of the forming gamete are not eliminated so they are transmitted to the gametes. This law fundamentally changes the concept (wrong all the way from Mendel and until now) regarding the content of the gametes provided by the parents to their descendants. The essence of this law consists in: in regular health conditions the gametes contain, aside the sex chromosome, only autosomal factors that were admitted in the set of active chromosomes (the factors that expressed and their latent reserves). This reality can be easily understood by carefully reading (Figure 1). |
| Â |
| Regarding the female gamete, it can be observed that the resemblance of the children more with the mother is manifested in case of participation at fecundation of each ovule (the chart would look the same for any ovule). So, both ovule-sister, formed in any meiosis, contain identical autosomal factors, the same that determined the mother’s phenotype, which can be found at the mother in the set of 22 active autosomal chromosomes. |
| Â |
| Regarding the male gamete on the chart it is noticed that it can exist and at the same time a resemblance of two children, girl and boy, more with the father. This means that both spermatozoon-brothers (the one with X and the one with Y), that determines the resemblance, can belong to the same lot, so they can be formed at the same time, meaning in the same meiosis. We assume, just like in the case of the ovules, that spermatozoon-brothers are gifted with identical factors, the ones in the set of 22 autosomal active chromosomes that determined the father’s phenotype. From the chart it can be assumed that the identical gifting is manifested to all spermatozoa from an individual, no matter the moment of their producing along the entire time of fertility of the male (the chart would look the same anytime). So, the following truth can be stated: both gametes-brothers, both at the male and at the female, receive, aside the sex chromosome, which is different, identical autosomal hereditary factors. In other words, any of the parents, father or mother, transmit by all their gametes, to all their descendants, the same autosomal factors, the same information; as a consequence, the factors in set of (n-1) inactive autosomal chromosomes are eliminated as factors Surplus, during meiosis; autosomal recessive diseases caused by deletions or duplicates are transmitted or not to the descendants, as well as the chromosome carrying the anomaly is chosen to be transmitted or not, according to the principle of the lowest energy consumption; from this law it can be assumed the fact that the crossing- over phenomenon, discovered by Morgan, is not a random process, like it was assumed until now, but one scheduled, coming from the necessity of separating active factors in order to be transmitted to the gametes and respectively inactive, in order to be eliminated. The way the law of gametes endowment works is easily understood from the (Figure 7). Because the complete (physically) autosome chromosomes obey the law the way they are, meaning that the active ones go into gametes and the inactive ones are eliminated, will be presented the case of combined chromosomes. |
| Â |
|
|
| Â |
| In meiosis, all factors that were admitted in the set of active chromosomes, at fecundation, are brought, by crossing-over, on only one chromosome of that certain pair. But, as on each of the two chromosomes of the pair (on the chart) are admitted factors (the ones marked with green line), one question arises: on each of the two chromosomes, from the father or from the mother, will be those factors brought? Answer: the universal principle, the lowest energy consumption, is applied once again. In this example they will be brought on the chromosome from the father (F); this is because, on the (F) chromosome there are 6 active factors, and on (M) chromosome there are only 4 (the ones with green line), the moving of the last ones obviously requiring a lower energy consumption. So, a crossing-over takes place between the factors of the pairs: G2, G4, G4, G5, the result being the two chromosomes in the right side of the figure, out of which the one with all the ex active factors will be included in the sets of the two gametes-brothers and the one with all the ex inactive factors will be eliminated. If it had been a reversed situation, on the (F) chromosome less active factors, and on (M) more, then all would have been brought on (M), then this would have been included in the gametes and (F) would have been eliminated. It can be observed on the chart, that both on (F) and on (M) there are also abnormal factors, L, D, i. These are not implied in the crossing-over process so each of them stays on its spot, being transmitted or not in the gametes, just like the carrying chromosome. For these factors, the crossing-over does not produce because: I have showed above that this process is scheduled to take place having a precise purpose and only among the factors that realised valid pairs, so capable to acquire the capacity of being active; at invalid pairs, not even a random crossing- over can be produced, as their factors are not eligible for the joining conditions (coupling), joining is the absolutely necessary phase that precedes the crossing-over, just like for acquiring the capacity of being active. [1,4,10,29-34]. |
| Â |
| Supplementary Information |
| Â |
| The supplementary informations for the article “Danciu laws of heredity - four new lawsâ€: 1 file (pdf 125 KB), “Supplementary Informationâ€, 9 figures and text - Applications that confirm the validity of the Danciu laws of heredity. |
| Â |
| Conclusion |
| Â |
| Danciu laws of heredity eliminate all errors of heredity made so far; completely clarifies the way characters are transmitted from parents to descendants, including most the autosomal - recessive diseases; are valid 100% in all cases and all species that reproduce by gametes; can have a significant practical importance in the creating activity of plants and animals hybrids with a high biological value; so, by a simple correlation of the moment of crosses/pairing with the appearance of the mature gametes, hybrids can be obtained with the most active hereditary factors from the most valuable individual; can be an important tool in the future research in genetics and mostly in the medical genetic research. |
| Â |
| |
| References |
| Â |
- Acharya A, Baek ST, Banfi S, Eskiocak B, Tallquist MD (2011) Efficient inducible Cre-mediated recombination in Tcf21 cell lineages in the heart and kidney. Genesis 49: 870-877.
- Ariel M, Robinson E, McCarrey JR, Cedar H (1995) Gamete-specific methylation correlates with imprinting of the murine Xist gene. Nat Genet 9: 312-315.
- Coimbra S, Salema R (1999) Ultrastructure of the developing and fertilized embryo sac of Amaranthus hypochondriacus L. Ann Bot 84: 781–789.
- Copeland NG, Jenkins NA, Court DL (2001) Recombineering: a powerful new tool for mouse functional genomics. Nat Rev Genet 2: 769-779.
- Doetschman T (2009) Influence of genetic background on genetically engineered mouse phenotypes. Methods Mol Biol 530: 423-433.
- Friedman WE (2001) Developmental and evolutionary hypotheses for the origin of double fertilization and endosperm. C R Acad Sci III 324: 559-567.
- Gehring M, Bubb KL, Henikoff S (2009) Extensive demethylation of repetitive elements during seed development underlies gene imprinting. Science 324: 1447-1451.
- Grossniklaus U, Vielle-Calzada JP, Hoeppner MA, Gagliano WB (1998) Maternal control of embryogenesis by MEDEA, a polycomb group gene in Arabidopsis. Science 280: 446-450.
- Guitton AE, Page DR, Chambrier P, Lionnet C, Faure JE, et al. (2004) Identification of new members of Fertilisation Independent Seed Polycomb Group pathway involved in the control of seed development in Arabidopsis thaliana. Development 131: 2971-2981.
- Nagy A (2000) Cre recombinase: the universal reagent for genome tailoring. Genesis 26: 99-109.
- Zhang Y, Riesterer C, Ayrall AM, Sablitzky F, Littlewood TD, et al. (1996) Inducible site-directed recombination in mouse embryonic stem cells. Nucleic Acids Res 24: 543-548.
- Agulnik AI, Mitchell MJ, Mattei MG, Borsani G, Avner PA, et al. (1994) A novel X gene with a widely transcribed Y-linked homologue escapes X-inactivation in mouse and human. Hum Mol Genet 3: 879-884.
- Barakat TS, Jonkers I, Monkhorst K, Gribnau J (2010) X-changing information on X inactivation. Exp Cell Res 316: 679-687.
- Basu R, Zhang LF (2011) X chromosome inactivation: a silence that needs to be broken. Genesis 49: 821-834.
- Beard C, Li E, Jaenisch R (1995) Loss of methylation activates Xist in somatic but not in embryonic cells. Genes Dev 9: 2325-2334.
- Borsani G, Tonlorenzi R, Simmler MC, Dandolo L, Arnaud D, et al. (1991) Characterization of a murine gene expressed from the inactive X chromosome. Nature 351: 325-329.
- Carrel L, Willard HF (2005) X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 434: 400-404.
- Carrel L, Park C, Tyekucheva S, Dunn J, Chiaromonte F, et al. (2006) Genomic environment predicts expression patterns on the human inactive X chromosome. PLoS Genet 2: e151.
- Chadwick BP, Valley CM, Willard HF (2001) Histone variant macroH2A contains two distinct macrochromatin domains capable of directing macroH2A to the inactive X chromosome. Nucleic Acids Res 29: 2699-2705.
- Chaumeil J, Waters PD, Koina E, Gilbert C, Robinson TJ, et al. (2011) Evolution from XIST-independent to XIST-controlled X-chromosome inactivation: epigenetic modifications in distantly related mammals. PLoS One 6: e19040.
- Cui S, Ross A, Stallings N, Parker KL, Capel B, et al. (2004) Disrupted gonadogenesis and male-to-female sex reversal in Pod1 knockout mice. Development 131: 4095-4105.
- Fang J, Chen T, Chadwick B, Li E, Zhang Y (2004) Ring1b-mediated H2A ubiquitination associates with inactive X chromosomes and is involved in initiation of X inactivation. J Biol Chem 279: 52812-52815.
- Huynh KD, Lee JT (2003) Inheritance of a pre-inactivated paternal X chromosome in early mouse embryos. Nature 426: 857-862.
- Lee JT (2000) Disruption of imprinted X inactivation by parent-of-origin effects at Tsix. Cell 103: 17-27.
- Baker P, Piven J, Schwartz S, Patil S (1994) Brief report: duplication of chromosome 15q11-13 in two individuals with autistic disorder. J Autism Dev Disord 24: 529-535.
- Christensen CA, Subramanian S, Drews GN (1998) Identification of gametophytic mutations affecting female gametophyte development in Arabidopsis. Dev Biol 202: 136-151.
- Maltais LJ, Lane PW, Beamer WG (1984) Anorexia, a recessive mutation causing starvation in preweanling mice. J Hered 75: 468-472.
- Liu Y, Yan Z, Chen N, Di X, Huang J, et al. (2010) Development and function of central cell in angiosperm female gametophyte. Genesis 48: 466-478.
- Baroux C, Spillane C, Grossniklaus U (2002) Evolutionary origins of the endosperm in flowering plants. Genome Biol 3: reviews1026.
- Cordts S, Bantin J, Wittich PE, Kranz E, Lörz H, et al. (2001) ZmES genes encode peptides with structural homology to defensins and are specifically expressed in the female gametophyte of maize. Plant J 25: 103-114.
- Gourdie RG, Green CR, Severs NJ, Anderson RH, Thompson RP (1993) Evidence for a distinct gap-junctional phenotype in ventricular conduction tissues of the developing and mature avian heart. Circ Res 72: 278-289.
- Hong SG, Oh HJ, Park JE, Kim MJ, Kim GA, et al. (2011) Production of offspring from cloned transgenic RFP female dogs and stable generational transmission of the RFP gene. Genesis 49: 835-840.
- Ingouff M, Jullien PE, Berger F (2006) The female gametophyte and the endosperm control cell proliferation and differentiation of the seed coat in Arabidopsis. Plant Cell 18: 3491-3501.
- Wolf CM, Berul CI (2006) Inherited conduction system abnormalities--one group of diseases, many genes. J Cardiovasc Electrophysiol 17: 446-455.
|
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â SUPPLEMENTARY INFORMATION |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â for |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â The Original Article |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Danciu Laws of Heredity |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â - four new laws - |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Author, Marin I. Danciu |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â e-mail: danciu@prodanciu.ro |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Applications that confirm |
| Â |
| Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â Â the Validity of the Danciu Laws of Heredity |
| Â |
| 1. The way of transmitting diseases caused by the non-function of the genes (factors) attached to X sex chromosome, like: Dalton disease (D), Haemophilia (H) etc. |
| Â |
|
|
| Â |
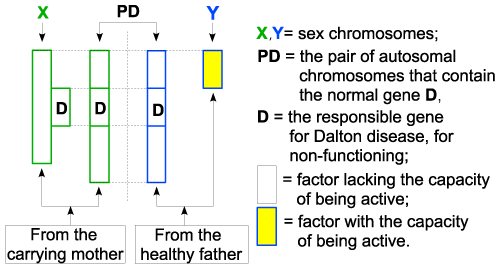
| Figure 1 – Applications |
| Â |
| I take for example a couple made of: |
| Â |
| - healthy father; |
| Â |
| - an apparently healthy mother, but carrier of an X chromosome of the gene (D)- all variants from 1 chromosome, which determines Dalton disease. |
| Â |
| The chart shows that the child will be a boy, by having XY chromosomes. The X chromosome originating from the mother, according to the law of sex chromosomes, does not have the capacity to be active in the boy’s body. As a result, the (D) gene on X does not have the capacity to be active, so being like the entire chromosome X. According to the autosomal hereditary factors law, the genes (D) on the two autosome chromosomes do not have the capacity to be active either, so they will capacitate one another during this fecundation, if the conditions mentioned in the law are fulfilled (only one pair of 2 genes at the same locus level, not identical). In this case, at fecundation, 3 genes and not only 2 so none of them will be capacitated. As a consequence, no gene (D) will be admitted in the set of active chromosomes (laws 1 and 3) which means that this gene will not be functional. The result will be a boy suffering from Dalton disease. |
| Â |
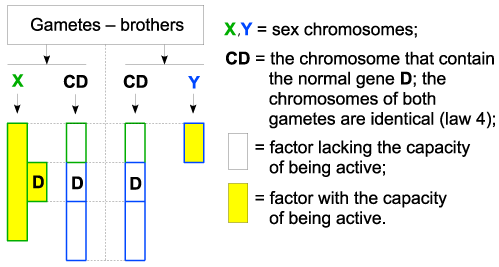
| At his turn, the boy will have the following gametes-brothers when reaching maturity (according to the law of gametes endowment): the chromosomes of both gametes are identical (law 4); |
| Â |
|
|
| Â |
| Figure 2 - Applications |
| Â |
| The children conceived with the participation of these gametes will be as follows: |
| Â |
| The boys |
| Â |
| The gamete that contains Y chromosome of the sick father and the gamete that contains X chromosome of a healthy mother, will participate at their conceiving, according to the following figure: |
| Â |
|
|
| Â |
| Figure 3 – Applications |
| Â |
| In this case, at fecundation, when meeting 2 regular genes (D), it is formed a valid pair, according to the law 3. As a result, the genes will mutual capacitate (law 3), one of them will be admitted in the set of active chromosomes (law 1), will be functional so the result will be a healthy boy. This result is in accordance with reality, the father suffering from Dalton disease will not transmit the disease to his boys. |
| Â |
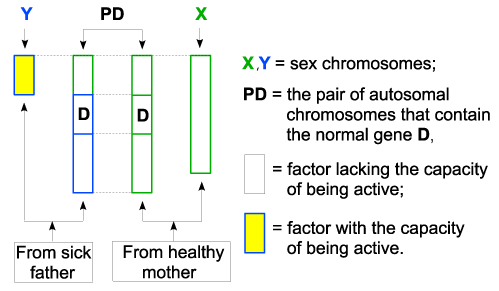
| The girls |
| Â |
| The gamete that contains X chromosome of the sick father and the gamete that contains X chromosome of a healthy mother, will participate at their conceiving, according to the following figure: |
| Â |
|
|
| Â |
| Figure 4 – Applications |
| Â |
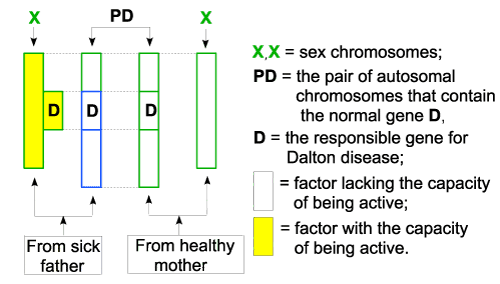
| In this case, the gene (D) attached to the X chromosome from the father already has the capacity to be active, because, according to the law 3 letter a), it was capacitated together with the X chromosome, also capacitated according to law 2. As a result, the two genes (D) on the pair of chromosomes, make a valid pair so they can join together and they can mutual capacitate, according to law 3. The gene (D) on X, having the valence for joining satisfied, cannot exert any influence on them. As a consequence, one of the capacitated genes (D) will be admitted in the set of active chromosomes (law 1 and 3), will be functional so, the girl will be apparently healthy. This result also reflects reality, so the girls born from fathers suffering from Dalton disease are just carriers of the disease, and apparently they are healthy. |
| Â |
| At their turn, these girls can have sick children, just like their grandmother, because of that X (D), but also healthy children, conceived with the participation of normal X chromosome (the one without D). |
| Â |
| Haemophilia and other disease caused by the non-functioning of the factor whose allele is attached to X chromosome are transmitted in the same way. I also highlight the fact that, the gravity of these diseases is different for each case: light forms, when the attached factors are of an inferior rank, meaning a small segment in a gene and severe forms, when factors of higher rank are attached to X, even an entire gene. |
| Â |
| 2. The way of transmitting some traits directly to the first generation or to the 2nd, the 3rd, etc. |
| Â |
| Let us take as trait the skin colour and the initial case as follows: |
| Â |
| - one partner is black (B); |
| Â |
| - the other partner is white (W). |
| Â |
| I also consider that the genes that encode the colour of the skin are 2 on each chromosome, so there are 2 variants (copies) with the same functional role, on each chromosome. |
| Â |
|
|
| Â |
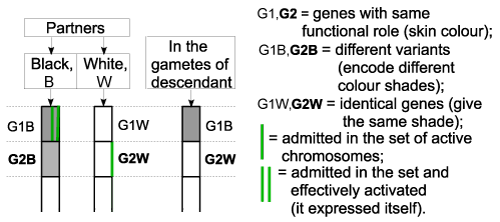
| Figure 5 - Applications |
| Â |
| In this example, right after fecundation, according to laws 1 and 3, the gene G1B (from black) and gene G2W (from white) were admitted in the set of active chromosomes. Then, it was marked the one that will be activated (law 3), only one of the 2 genes with the same functional role, more precisely the gene G1B. The result is that the child’s skin will be black, determined by the expression of the gene G1B. But its gametes will contain (law 4), aside the gene G1B, also the gene G2W, because this one was also admitted in the set of active chromosomes, where it remained latent. |
| Â |
| It can be noticed on the chart, that genes G1W and G2B were eliminated (in meiosis), according to law 4, so these were not transmitted to the descendant. |
| Â |
| The gametes of the descendant contain the gene G2W also, for the white colour, although its body has a black colour. |
| Â |
| If gene G1, chosen to express itself, would be a combined gene (law 1), meaning that it would have a segment on G1B and the complementary segment on G1W, the skin colour should be a mixture of black and white (mulatto). |
| Â |
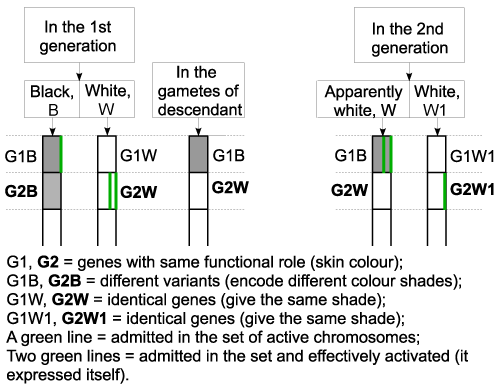
| I will illustrate you, using the same example, the transmitting of the white colour to the first generation, then the transmitting of the black colour to the 2nd generation. (Figure 6 - Applications) |
| Â |
| In the 1st generation |
| Â |
| It is noticed on the chart that gene G2W will express itself so the child’s skin will be white. Its gametes will contain, aside the gene G2W, the gene G1B, which encoded the black skin. |
| Â |
| In the 2nd generation |
| Â |
| The apparently white individual, resulted in the 1st generation, but carrier of the gene for black skin, together with a complete white individual, can have children with black skin in the second generation, according to the chart in the right side of the figure. G1B is activated here, meaning that the child will have black skin, although, apparently, none of his parents is black. |
| Â |
|
|
| Â |
| Figure 6 - Applications |
| Â |
| I will illustrate the combination between two gametes resulted in the 1st generation from the two couples, each of them consisting of a black and a white partner. |
| Â |
|
|
| Â |
| Figure 7 – Applications |
| Â |
| But, each child will have the two genes in the set of active chromosomes: one that expresses itself and determines the apparently colour of the skin and the other one latent but which encodes the other colour. |
| Â |
| 3.Other examples that confirms the possibility of increasing and decreasing the number of individuals in the cases when the partners are relatives. |
| Â |
| I assume that instead of 2 variants on the same chromosome, like we had before, now there are 5 variants of the gene that encodes a certain trait noted with G1, G2, G3, G4, G5. |
| Â |
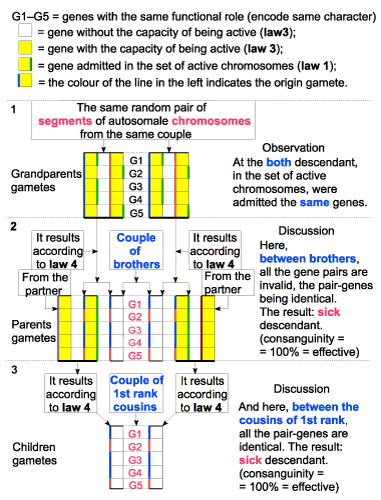
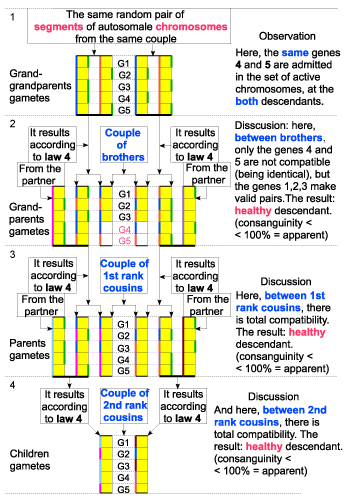
| I have taken into consideration a couple that has two descendants of different gender, then, each of them has other descendants (Figure 8 - A and Figure 9 - B). |
| Â |
| Conclusions for the cases in which the partners are relatives |
| Â |
| 1. By the above mentioned charts, when applying the new laws of heredity, it is confirmed the fact that both possibilities exist indeed: the result can be either healthy descendants, or sick ones, so the number of individuals can be either increasing or decreasing, depending on the proportion of consanguinity. |
| Â |
| 2. As the number of genes (variants) with same functional role (on the same chromosome) is bigger, the risk of appearing sick descendants is smaller. |
| Â |
| 3. So, we can assume that this number varies from one species to another and it is likely to be bigger a species that use to live in groups. |
| Â |
| Results when partners are relatives |
| Â |
|
|
| Â |
| Figure 8 - A. The descendants that resulted are sick |
| Â |
|
|
| Â |
| Figure 9 - B. The descendants are healthy |