Biostratigraphic Revision of Middle-Cretaceous Succession in South Zagros Basin (SW of Iran)
Received: 20-May-2014 / Accepted Date: 21-Aug-2014 / Published Date: 30-Aug-2014 DOI: 10.4172/2157-7617.1000216
Abstract
Cenomanian succession (Sarvak Formation) is well distributed along Zagros area particularly in the south portion of the Zagros (Fars area). This study attempts to determine the precise biostratigraphic biozones of four stratigraphic sections selected from the Cenomanian strata south Zagros basin, including the sections of Nour Abad, Samghan, Doroodzan, and Arsanjan. The vertical distribution of the index foraminifers confirms a faunal assemblage of benthic and planktic foraminifers in the mentioned stratigraphic sections. The received Biostratigraphic data of this work reveals well developed benthic foraminifers and well distributed Cenomanian planktons in the uppermost lithostratigraphic part of the studied Sarvak exposures. However, the established biozones of the Sarvak Formation represent the Cenomanian age, which is similar to many recognized Cenomanian biozones in the Tethyan realm, whereas the foraminiferal biostratigraphy of the Arsenjan stratigraphic column is assigned to the Late Albian to Early Cenomanian age. Since benthic foraminifers are well expanded in studied section of Sarvak Formation, the presence of pelagic biofacies is a proof of transgression in some studied sections. However, the established biozones support that geologic setting is the most factor which controls vertical distribution of foraminifers and other microfossils during the deposition of Sarvak Formation. The received biostratigraphic data of the Sarvak Formation determines subdivision of Cenomanian age in the South Zagros basin clearly.
Keywords: Biostratigraphy; Foraminifera; Cenomanian; Zagros; Iran
6330Introduction
Many geologists have focused upon the carbonate petroleum reservoir rock of the Zagros area because of the well-known oil productivity of this region [1]. One of the most oil producing reservoir rocks of the Zagros basin is the Asmari Formation (Oligo-Miocene) which is well known for petroleum geologists [2]. In order to develop oil fields in the Zagros mountain ranges, the National Iranian Oil Company (NIOC) established a progressive study of other carbonate facies in the Zagros area. The Sarvak Formation is one of the most significant carbonate reservoir rocks known to be an oil producer in the Zagros oil field [3]. Oil drilling projects reveals Sarvak Formation as the most important reservoir rocks in south Zagros Basin [3,4].
Actually, the Sarvak Formation is a thick carbonate lithostratigraphic unit of the Zagros located on the southern border of Neotethys. This rock unit is related to the Bangestan group in the Zagros region. The Mid-Cretaceous succession is a sedimentary cycle which consists of rock units similar to that of the Sarvak, Surgah and Ilam as Bangistan group. Since Sarvak Formation has been determined to be one of the most important reservoir rocks in the Zagros basinthe biostratigraphy study proves to be of significance in the targeting of oil field development for Iranian petroleum geologists. The stratigraphic position of the Sarvak Formation is also another reason for the importance of the biostratigraphic study. According to James and Wynd [5], the type section of the Sarvak Formation was selected in the Bangestan anticline in Khuzestan province (SW of Iran). The age determination of Sarvak was assigned to the Albian through to the Cenomanian ages. Based on the study, Zagros is divided into four main zones: Khuzestan, Lurestan, Coastal Fars and Interior Fars. Ghavidel Syooki et al. [6] have documented valuable biostratigraphic data from the Paleozoic sequence of the Zagros area (Figure 1). The fundamental biostratigraphic study of the Zagros basin was provided by Wynd [7]. He suggested that Cenomanian benthic foraminifers are well expanded in the Khuzestan, Coastal and Interior Fars areas whereas pelagic facies are well dominant in the Lurestan region. In order to determine the lower and upper lithostratigraphic boundary of the Sarvak Formation, Motiei [8] considered the surface and subsurface stratigraphic data which have been obtained from the exploration section of the National Iranian Oil Company. Based on the study, the Sarvak Formation covers the Kazhdumi Formation along the Zagros Basin; the upper lithostratigraphic limit of Sarvak limestone is marked by sharp contact between the Sarvak and Ilam Formations in Khuseztan and some parts of Coastal Fars regions. Likewise, Afghah and Giyasi [9] have reported that the Gurpi Formation (Late Campanian–Maastrichtian) is an overlaying formation in some parts of Interior Fars area with obvious disconformity. Since the lithological aspect of Kazhdumi is comprised of shaly and marly limestones, the lower lithostratigraphy of the Sarvak Formation is marked by sharp contact.
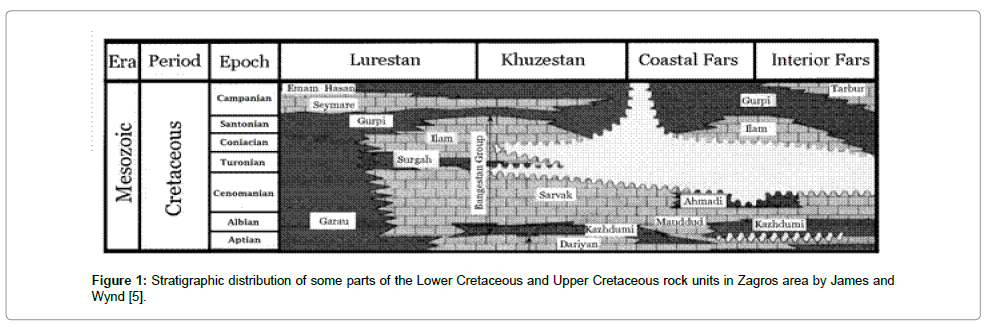
Figure 1: Stratigraphic distribution of some parts of the Lower Cretaceous and Upper Cretaceous rock units in Zagros area by James and Wynd [5].
Kalantari [4] has recorded the disconformity between the Sarvak and Ilam Formations near Shiraz city (SW Iran) previously. Because of the importance of the Sarvak Formation as reservoir rocks, many researchers have studied it [10-12]. However the biostratigraphic significance of the Sarvak Formation is applicable in oil drilling projects, particularly in Fars area.
Materials and Methods
In order to conduct a biostratigraphic study of Mid-Cretaceous strata in the south Zagros region, four stratigraphic sections of the Sarvak Formation were selected, which are named: Nour Abad, Doroodzan, Samghan and Arsenjan. The upper and lower lithostratigraphic limits were determined by detailed field work. Based on field observations, lithological aspects were distinguished and the thickness of both stratigraphic sections was measured. In addition, 440 samples were collected systematically from the four studied sections and coded separately. The biostratigraphy method is based on study of stratigraphic distribution of benthic, planktic foraminifers and other microfossils. However, the mentioned method is significant in all biostratigraphy of Cretaceous strata in the Zagros basin. According to the mentioned method, thin sections were provided from the collected samples and studied in detail. Microscopic figures were prepared at the Geology Department of the Islamic Azad University of Shiraz. Since the biofacies of both studied sections are mainly comprised of benthic and pelagic foraminifers, the identification of Mid-Cretaceous foraminifers was provided [13-24]. The vertical distribution of identified formainifers and other microfossils led us to established biozones. Eventually, described biozones were campared to previous Zagros studies and other biostratigraphy works of Tethyan realm.
Geographical and Geological Setting
The study area is located between E: 51º31' to E: 53º04' and N: 21º52' to N: 30º13' (Figure 2) south Zagros Basin. Four stratigraphic sections of the Sarvak Formation were selected for this work. The studied sections nominated were Nour Abad, Doroodzan, Samghan and Arsenjan. The detailed geographic coordinates of each section are as follows: E: 51º30' 12" and N: 30º08' 20" (Nour Abad), E: 52º26' 43" and N: 30º12' 31" (Doroodzan); E: 51º40'17" and N: 21º 52' 50" (Samghan); E: 53º 03' 87" and N: 29º 52' 34" (Arsenjan). The Nour Abad, Doroodzan and Samghan sections are located in the northwest of Shiraz and the locality of the Arsenjan section is in northeast Shiraz. According to Alavi [25], both four study sections are located in a simple folded zone of the Zagros. Actually, the simple folded zone consists of many simple box folds which are oriented in a northwest–southeast trend. The Nour abad stratigraphic section is located in a well-known faulted zone of the south Zagors Basin. The most characteristic of this zone is assigned to normal stratigraphic position of the Zagros rock units, which has been confirmed by many workers [25-27]. Based on James and Wynd [5], the Arsenjan and Doroodzan stratigraphic sections are located in the Interior Fars area, whereas the Nour Abad and Samghan sections are located in the Coastal Fars region.
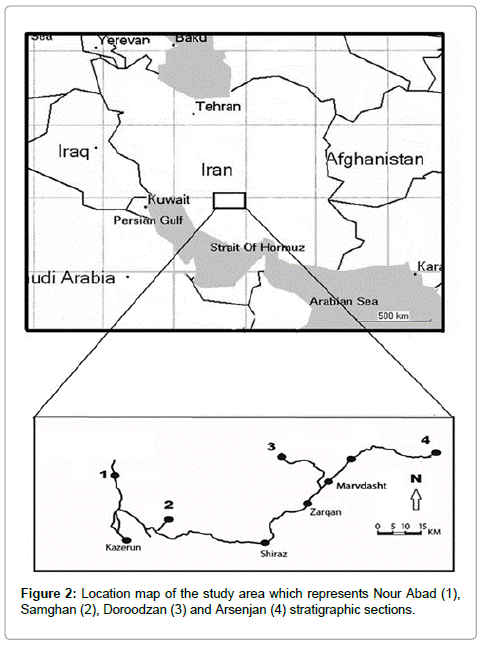
Figure 2: Location map of the study area which represents Nour Abad (1), Samghan (2), Doroodzan (3) and Arsenjan (4) stratigraphic sections.
Stratigraphy
The Sarvak Formation consists mainly of grey limestone which is sandwiched between the Kazhdumi and Gurpi Formations in the Nour Abad area. The detailed field work on the studied stratigraphic sections shows the thickness of the Sarvak outcrop measures about 345 meters. The lower and upper lithostratigraphic limits of the Sarvak Formation are well exposed in the mentioned area. The lower contact of the Sarvak limestone is marked by a sharp lithologic change from grey Sarvak limestone to green marls of the Kazhdumi formation. The light grey shale of the Gurpi Formation covers the Sarvak Formation disconformably as a sharp contact. The Sarvak exposure is made of thick to medium-bedded grey limestone and dark grey massive limestone in the Nour Abad region. Microscopic investigations have confirmed the presence of well dominant bioclast wackestone, rudist packstone, foraminifer intraclast wackestone and Calcisphaerula innominata wackestone as major lithofacies of the Sarvak Formation studied in the Nour Abad section.
The Sarvak Formation in the Arsenjan stratigraphic column is extended by approximately 105 meters. The lower lithostratigraphic limit of the Sarvak limestone covers a greenish-grey thin- to mediumbedded marly limestone of the Kazhdumi Formation as a sharp contact. Similar to the Nour Abad area, the Kazhdumi Formation consists of green to grey thin- to medium-bedded Orbitolinid yield as well as abundant planktons distinguished in the marly limestone of the middle to upper portion of this rock unit. The mentioned marls are very rich in iron nodules, particularly, in lower lithostratigraphic limit of the Kazhdumi formation. The upper contact of the Sarvak is not clear because of the erosional processes in the study area. The Sarvak Formation is comprised of grey thick- to medium-bedded limestone with argillaceous marls as interbeds; the argillaceous limestone is also well expanded in the upper part of this stratigraphic section. Generally, pelagic facies are well distributed along the Sarvak stratigraphic section of Arsenjan. Crushed rudist wackestone, Calcisphaerula innominata and rudist fragment wackestone are common in the lithofacies of the Sarvak Formation. Sarvak outcrops of the Samghan area measure 204 meters, where thick massive grey limestone alternates with grey medium-bedded limestone. The lower contact of the Sarvak limestone covers marls of the Kazhdumi Formation with a vertical change in lithology. Gurpi shale is overlaid with Sarvak limestone at a distinct lithostratigraphic change, which is marked by an erosional surface and iron nodules. The Sarvak Formation of the Doroodzan stratigraphic section measures approximately 207 meters, which includes a grey thick to massive limestone with iron nodules that alternates with dark grey thin to medium limestone. It terminates with dark gray massive limestone with Exogyra sp. and rudist fragment beds. Other similar studied stratigraphic sections of the lower lithostratigraphic limit of the Sarvak Formation cover marls and marly limestone of the Kazhdumi Formation whereas the Ilam formation is overlaid on the upper lithostratigraphic limit of the Sarvak limestone in this section disconformably.
Biostratigraphy
The vertical distribution of identified species and genus of the stratigraphic study of the Sarvak Formation supports a different biozonation of each section. Actually, the different paleoenviornmental conditions cause various biozones in each studied stratigraphic section. The established biozones comprise benthic and planktonic microfossils. Foraminifers are valuable in the biostratigraphic study of Sarvak sections as they are used as the main tools for the establishment of biozones. Detailed biostratigraphical study led us to establish the biozones of the Sarvak Formation as follows:
Arsanjan biostratigraphy
According to microscopic investigations of the Arsenjan stratigraphic section, one biozone named the Calcisphaerula innominata and Assemblage Zone was established (Figure 3). Based on identified foraminifers, eleven species were confirmed which are common and well distributed in this stratigraphic section.
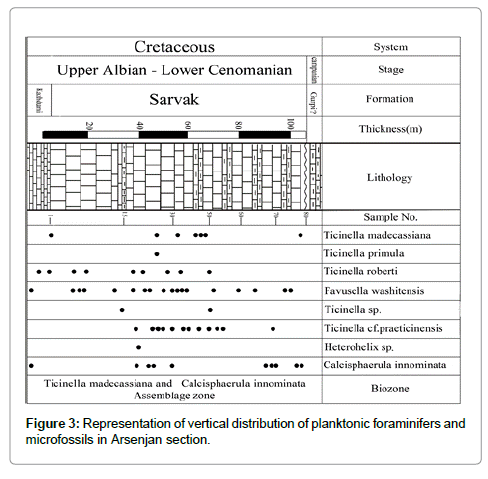
Figure 3: Representation of vertical distribution of planktonic foraminifers and microfossils in Arsenjan section.
The Ticinella madecassiana and Calcisphaerula innominata Assemblage Biozone is described by the first occurrence of Calcisphaerula innominata Bonet and Ticinell madecassiana Sigal. The mentioned biozone is made up of 105 metres of grey medium- to thick-bedded limestone; actually, it is made of Calcisphaerula bioclast wackestone to packstone sublithofacies. The microscopic study of the Sarvak lithofacies in this section reveals the crushed angular rudist fragments with pelagic foraminifer species and the very dominant Calcisphaerula innominata Benot. The lower biostratigraphic of this biozone is marked by the last occurrence of Hemicyclammina sigali Maync, which is distinguished in the uppermost lithostratigraphic part of the Kazhdumi Formation. The faunal assemblage which is associated with Ticinella madecassiana Sigal and Calcisphaerula innominata Bonet are: Ticinella roberti (Gandolfi), Ticinella cf. praeticinensis Sigal, Ticinella primula Luterbucher, Heterohelix sp., and Favusella washitensis (Carsey). Permoli Silva and Verga [16] have documented Ticinella madecassiana Sigal from Upper Albian through to the Lower Cenomanian strata of the Tethyan realm. According to their work, Ticinella roberti (Gandolfi) and Ticinella praeticinensis Sigal are assigned to the Lower Albian–Lower Cenomanian strata. Khosrow Tehrani and Fonooni [10] have documented Favusella washitensis (Carsey) from Upper Albian–Lower Cenomanian strata of the Bangestan anticline (SW of Iran). Therefore, the Upper Albian–Lower Cenomanian age of this biozone is confirmed by referring to diagnosed faunal assemblage. Likewise the range zone of Ticinella madecassiana Sigal, Ticinella primula is documented from Middle Albian to Lower Cenomanian. Husinec et al. [13] have reported Hemicyclammina sigali Maync from the Lower Albian sequence of the west of Croatia. Ghanem and Kuss [28] have documented this taxon from the Early to Late Albian sediments of northwest Syria. It should be noted that there are many records which reflect different stratigraphic ranges of Hemicyclammina sigali Maync [29,30]. Since the last occurrence of Hemicyclammina sigali Maync is synchronous with the first presence of Ticinella medecassiana Sigal and Calcisphaerula innominata Benot in this section, it confirms that this biozone refers to the Late Albian– Early Cenomanian age, which is comparable with biozone number 23 of Wynd [7].
Biostratigraphy
The vertical distribution of identified species and genus of the stratigraphic study of the Sarvak Formation supports a different biozonation of each section. Actually, the different paleoenviornmental conditions cause various biozones in each studied stratigraphic section. The established biozones comprise benthic and planktonic microfossils. Foraminifers are valuable in the biostratigraphic study of Sarvak sections as they are used as the main tools for the establishment of biozones. Detailed biostratigraphical study led us to establish the biozones of the Sarvak Formation as follows:
Arsanjan biostratigraphy
According to microscopic investigations of the Arsenjan stratigraphic section, one biozone named the Calcisphaerula innominata and Assemblage Zone was established (Figure 3). Based on identified foraminifers, eleven species were confirmed which are common and well distributed in this stratigraphic section.
The Ticinella madecassiana and Calcisphaerula innominata Assemblage Biozone is described by the first occurrence of Calcisphaerula innominata Bonet and Ticinell madecassiana Sigal. The mentioned biozone is made up of 105 metres of grey medium- to thick-bedded limestone; actually, it is made of Calcisphaerula bioclast wackestone to packstone sublithofacies. The microscopic study of the Sarvak lithofacies in this section reveals the crushed angular rudist fragments with pelagic foraminifer species and the very dominant Calcisphaerula innominata Benot. The lower biostratigraphic of this biozone is marked by the last occurrence of Hemicyclammina sigali Maync, which is distinguished in the uppermost lithostratigraphic part of the Kazhdumi Formation. The faunal assemblage which is associated with Ticinella madecassiana Sigal and Calcisphaerula innominata Bonet are: Ticinella roberti (Gandolfi), Ticinella cf. praeticinensis Sigal, Ticinella primula Luterbucher, Heterohelix sp., and Favusella washitensis (Carsey). Permoli Silva and Verga [16] have documented Ticinella madecassiana Sigal from Upper Albian through to the Lower Cenomanian strata of the Tethyan realm. According to their work, Ticinella roberti (Gandolfi) and Ticinella praeticinensis Sigal are assigned to the Lower Albian–Lower Cenomanian strata. Khosrow Tehrani and Fonooni [10] have documented Favusella washitensis (Carsey) from Upper Albian–Lower Cenomanian strata of the Bangestan anticline (SW of Iran). Therefore, the Upper Albian–Lower Cenomanian age of this biozone is confirmed by referring to diagnosed faunal assemblage. Likewise the range zone of Ticinella madecassiana Sigal, Ticinella primula is documented from Middle Albian to Lower Cenomanian. Husinec et al. [13] have reported Hemicyclammina sigali Maync from the Lower Albian sequence of the west of Croatia. Ghanem and Kuss [28] have documented this taxon from the Early to Late Albian sediments of northwest Syria. It should be noted that there are many records which reflect different stratigraphic ranges of Hemicyclammina sigali Maync [29,30]. Since the last occurrence of Hemicyclammina sigali Maync is synchronous with the first presence of Ticinella medecassiana Sigal and Calcisphaerula innominata Benot in this section, it confirms that this biozone refers to the Late Albian– Early Cenomanian age, which is comparable with biozone number 23 of Wynd [7].
Nour abad biostratigraphy
The biostratigraphy of the Nour abad Sarvak Formation indicates two species of plankton and two genera and 15 species of benthic foraminifers. Based on the stratigraphic distribution of diagnosed foraminifers, two biozones are established: (1) The Conicorbitolina conica Taxon Range Zone, and (2) the Nezzazata simplex and Chrysalidina sp. Assemblage Zone (Figure 4).

Figure 4: Stratigraphic distribution of foraminifers in Nour Abad section.
This biozone is distinguished in the lower portion of this stratigraphic section by the first representatives of the species Conicorbitolina conicaD´Archiac. The lithological aspect of this biozone consists of dark grey medium-bedded limestone. This biozone is overlaid on the Kazhdumi formation, which extends for about 70 meters. Microscope investigations present the alternation of bioclastic wackestone, Orbitolina wackestone and Orbitolina grainstone as major sublithofacies of this biozone. Rarely bioclastic packstone is distinguished. Diagnosed foraminifers of this biozone include Nezzazata sp. and Sellialveolina vialli Colalongo. Calonge et al. [31] reported that Sellialveolina vialli Colalongo has also been investigated in the lowermost of the Cenomanian Iberian Range (Spain). Bosellini et al. [32] have also recorded Sellialveolina vialli Colalongo from the Early Cenomanian strata of Masseria Quadrone Limestone (east of Italy). Sartorio et al. [33], Simmons et al. [34] and Tertis et al. [35] have confirmed the Lower Cenomanian age for this faunal assemblage. According to Simmons et al. [34] and Vilas et al. [36], the Orbitolinid yield from strata illustrates the dominance of the shallow water carbonate paleoenvionment in this biozone. Based on the identified faunal assemblage, this biozone was assigned to the Early Cenomanian age. This zone is comparable to biozone number 23 of Wynd [7].
Biozone 2: (Nezzazata conica and Chrysalidina sp. Assemblage Zone)
The lithologic aspect of this biozone is comprised of light to dark grey medium- to thick-bedded limestone which measures approximately 120 meters in thickness. Identified sublithofacies of this biozone include intraclast bioclast grainstone and bioclast wackestone. Similar to the Conicorbitolina conica Taxon Range Zone, bioclastic packstone is recognized rarely.
The biozone Nezzazata conica Smouth and Chrysalidina sp. is recognizable in the middle portion of this section which is marked by the first appearance of Nezzazata conica Smout and Chrysalidina sp. The last occurrence of the mentioned foraminifer taxa is synchronous with the first appearance of Nummoloculina heimi Bonet and Pseudorhipidionina casertana DE Castro. Common foraminifers of this biozone consist of Nezzazata simplex Omara, Nezzazata sp., Nezzazatinella picardi Henson and Biconcava bentori Hamaoui and Saint-Marc. Aguilar-Franco [15] has documented Nezzazata conica Smout from the Cenomanian sequence of southern Mexico. It should be noted that the Mid-Cenomanian boundaries are not clear in the mentioned area, whereas Husinec et al. [13] have investigated this taxon from Middle Cenomanian strata of the Baldarin stratigraphic section which is located at Cres Island (Croatia). Likewise, they have investigated Nezzazata simplex Omara, Nezzazatinella picardi Henson and Psudolituonella reicheli Marie from the Mid-Cenomanian of the mentioned stratigraphic section. Other biostratigraphic data confirm the vertical distribution of this taxon (Nezzazata conica Smout) from Early–Middle Cenomanian sediments of the Adriatic platform, Croatia [14]. Another record of the stratigraphic distribution of this taxon reveals the Late Cenomanian age of the central Italy succession [19]. It should be noted that he has not marked the Mid-Cenomanian in his work. Recognition of Nezzazata conica Smout has been confirmed by Sari et al. [37] in the Middle to Upper Cenomanian sequence in the southwestern area of Turkey. However, Nezzazata simplex Omara has been recorded as a wide ranging taxon from the Early Cenomanian through to Late Campanian of the Karst Dinarides region (SE Europe) by Velic [18]. Various recordings of the Nezzazata simplex taxon range zone have been documented by Sari et al. [37], which have been reflected in the Mid- to Late-Cenomanian strata of SW Turkey. Biozone 2 has a similar characteristic value to Wynd´s [7] biozone number 25. However, many documents have confirmed the Middle Cenomanian age of this foraminiferal association [38,39]. Therefore, it is confirming the Middle Cenomanian age of this biozone.
Biozone 3: (Nummoloculina heimi zone)
This biozone is recognized in the upper part of this stratigraphic section, which is investigated by the first occurrence of Nummoloculina heimi Bonet. This biozone is made of grey massive limestone which is composed alternately of intraclast wackestone, pell bioclast wackestone and interaclast bioclast grainstone. The thickness of this biozone is measured at approximately 90 meters. The investigated foraminifer associations of this biozone include: Pseudorhipidionina casertana DE Castro, Psudolituonella reicheli Marie, Praealveolina cretacea (D’Archiac) and Cuneolina pavonia D’Orbigny which are distinguishable in bioclastic packstone and grainstone lithofacies. Hernández-Romano et al. [40] reported Nummoloculina heimi Benot from the Late Cenomanian sequence of southern Mexico. Likewise similar data were obtained by Borghi and Pignatti [41] from Upper Cenomanian sediments in southern Italy. Furthermore, other biostratigraphic studies have confirmed the Upper Cenomanian age of Nummoloculina heimi Benot [40,41]. Cuneolina pavonia D’Orbigny has been reported from the Albian to Cenomanian succession of Tethys by Filkorn and Scott [42]. This species is recorded from Middle Cenomanian of west central Sinai (Egypt) by Orabi et al. [23] whereas Praealveolina cretacea (D’Archiac) has been distinguished as the Praealveolina cretacea zone, which indicates Late Cenomanian age in the mentioned area. However, the stratigraphic position of this zone is assigned to Late Cenomanian age. The faunal assemblage of this biozone is similar to biozone numbers 24 and 25 of Wynd´s biozonation [7].
Biozones 4: (Calcisphaerula innominata and Whiteinella paradubia Assemblage Zone).
This biozone extends in the upper portion of this stratigraphic section. The first appearance of Calcisphaerula innominata Benot and Whiteinella paradubia (Sigal) is an indication of this biozone which is overlaid on Biozone 3. This biozone is characterized by dark grey limestone. The major identified lithofacies of this biozone is Calcisphaerula wackestone, which consists of Late Cenomanian pelagic foraminifers. It measured 65 meters and is underlaid by Gurpi shale disconformably. Phitonella ovalis Kaufm is associated with mentioned microfossil taxa. These taxa are distinguished remarkably well in the Calcisphaerula wackestone. Permoli Silva and Verga [16] have marked Whiteinella paradubia (Sigal) from Mid-Cenomanian through to Mid-Coniacian, whereas Keller and Pardo [43] have reported this species from the Upper Cenomanian strata of the Greenhorn formation of Colorado (USA). The biostratigraphic results of Cetean et al. [44] are similar to the work of Permoli Silva and Verga [16] who studied the eastern Carpathians. Based on the mentioned work, it confirmed the Late Cenomanian age of biozone Calcisphaerula innominata and Whiteinella paradubia Assemblage Zone. The characteristic values of this biozone are close to those of biozone number 26 of Wynd [7].
Samghan biostratigraphy
The lower limit of the Sarvak Formation with the Kazhdumi Formation is gradational and conformable, formed of thick-bedded gray limestone; its upper lithostratigraphic limit is disconformable with pelagic shale of the Gurpi Formation. Measured thickness of the Sarvak Formation in this section is 204 meters and it includes light to dark gray medium- to thick-bedded limestone. It terminates in massive gray limestone with iron nodules. Based on foraminiferal studies, two assemblage biozones have been suggested for this section. Established biozones include the following: Orbitolina sp. Conicorbitolina conica Assemblage Zone; Nezzazata concava and Parealveolina cretacea- Assemblage Zone; and Pseudolituonella reicheli, Pseudorhipidionina sp. and Chrysalidina gradata Assemblage Zone (Figure 5).

Figure 5: Illustration of established biozones and faunal assemblages in Samghan section.
Biozone 1: (Orbitolina sp., Conicorbitolina conicaand Dictyoconus sp. Assemblage Zone)
This biozone is characterized by grey medium-bedded limestone which extends about 32 meters. However, Orbitolina sp., Conicorbitolina conica, and Dictyoconus sp. biozone is composed of peloid skeletal wackestone to packstone, which confirms shallow water, carbonate system during deposition. The first occurrence of Orbitolina sp., Conicorbitolina conica(D'Archiac) and Dicyoconus sp. marks this biozone. The faunal assemblage of this zone consists of: Ovalveolina ovum D’Orbigny, Ovalveolina crassa DE Castro, Ovalveolina sp., Praealveolina tenuis Reichel, Nezzazata conica Smouth, Nezzazatinella picardi (Henson). Aguilera-Franco [15] documented some taxa with a wide range from southern Mexico (e.g. Nezzazata conica Smouth and Nezzazatinella picardi (Henson). Velic [18] distinguished similar biostratigraphic data from Karst Dinarides (SE Europe). Husinec et al. [13] obtained a stratigraphic distribution of Orbitolinids from western Croatia by Fucek et al. [45]. Vertical distribution of Orbitolinids confirms that Conicorbitolina conica(D'Archiac) is the first occurrence from the Lower Cenomanian strata of mentioned area. Other workers have documented similar data [13,45] from western Croatia. Husinec et al. [14] reported Ovalveolina crassa DE Castro from the Early Cenomanian sequence of the Adriatic platform (western Croatia). Based on their work other species have been investigated with Ovalveolina crassa DE Castro (e.g. Conicorbitolina conica (D'Archiac) and Sellialveolina viallii). This taxon has been recognized with Praealveolina simplex Pichel, Pseudocyclammina rogusa D’Orbigny of biozone no.1 (Early Cenomanian) in Karst Dinarides [18]. Ovalveolina ovum D’Orbigny has been recognized from the Middle to Upper Cenomanian sediments [46-48]. The first occurrence of this taxon has been documented from Lower Cenomanian of the Sarvestan succession (SW Iran) by Kalantari [4]. Additionally, the identified foraminifers confirm the Early Cenomanian age of this biozone (Figure 6).
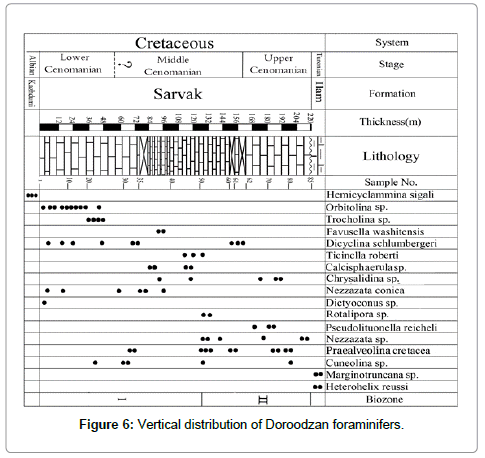
Figure 6: Vertical distribution of Doroodzan foraminifers.
Biozone 2: (Nezzazata concava and Praealveolina cretacea Assemblage Zone)
This is lithologically composed of thick to massive gray limestone with grey medium-bedded limestone which consists of iron nodules. This biozone extends approximately 78 meters. Major identified sublithofacies are comprised of foraminifer wackestone, foraminifer bioclast wackestone to packstone and intraclast bioclast packstone. Generally, common bioclasts are identified as crushed bivalves.
This biozone is distinguished by the first occurrence of both Nezzazata concava (Smout) and Praealveolina cretacea (D’Archiac). Diagnosed foraminifers of this biozone include: Simplalveolina simplex Reichel, Ovalveolina ovum D’Orbigny, Dicyclina schlumbergeri Munier-Chalmas, Dictyoconus pachymarginalis Schroeder, Cuneolina sp., Nezzazata simplex Omara, Nezzazata sp., Choffatella sp., Biplanata peneropliformis Hamaoui and Saint-Marc, Nezzaztinella picardi (Henson), Nummoloculina regularis Philipson, Cisalveolina frassi (Gumbel), Cycledomia iranica (Henson) and Trochospira avnimelechi Hamaoui and Saint-Marc. Hart et al. [48] have suggested Simplalveolina simplex Reichel is recognizable in the Middle to Upper Cenomanian faunal assemblage of shallow water-rich environments. Husinec et al. [14] recorded Nezzazata simplex Omara from the Upper Albian through to Middle Cenomanian sequence of the Adriatic platform. Likewise similar biostratigraphic data have been documented from the Baldrin section of Croatia [13]. Different ranges of this taxon have been investigated by many workers [15,18,23,40] represented Nezzazata concava (Smout) as an association of Upper Cenomanian sediments in west-central Sinai (Egypt). Based on their work, Nezzazata concava (Smout) is recognizable in the Praealeveolina cretacea zone. According to Ghanem et al. [28], the first appearance of Praealveolina cretacea (D’Archiac) is documented from the Early to Mid Cenomanian of Syria. Likewise biostratigraphic data on Nezzazata concava (Smout) are acceptable proof for the assignment of Late Cenomanian age to this biozone (Figure 7).
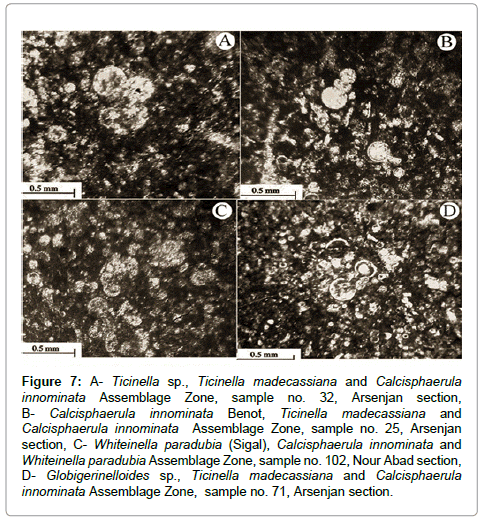
Figure 7: A- Ticinella sp., Ticinella madecassiana and Calcisphaerula innominata Assemblage Zone, sample no. 32, Arsenjan section, B- Calcisphaerula innominata Benot, Ticinella madecassiana and Calcisphaerula innominata Assemblage Zone, sample no. 25, Arsenjan section, C- Whiteinella paradubia (Sigal), Calcisphaerula innominata and Whiteinella paradubia Assemblage Zone, sample no. 102, Nour Abad section, D- Globigerinelloides sp., Ticinella madecassiana and Calcisphaerula innominata Assemblage Zone, sample no. 71, Arsenjan section.
Biozone 3: (Pseudolituonella reicheli, Pseudorhipidionina sp., and Chrysalidina gradata Assemblage zone)
The lithologic characteristics of this biozone mainly consist of light to dark grey medium- to thick-bedded rudist limestone with macrofossil debris, particularly Exogyra sp. This biozone has been measured as about 106 meters in thickness. Detailed microscopic study reveals the alternation of bioclastic wackestone with packstone as the major identified sublithofacies of this biozone. This biozone is marked by the first occurrence of Pseudolituonella reicheli Marie, Pseudorhipidionina sp. and Chrysalidina gradata D'Orbigny. The upper biostratigraphic limit of this biozone is distinguishable by the recognition of Campanian planktons of the Gurpi formation. Common foraminifers of this biozone include: Dictyoconus arabicus Henson, Dictyoconus pachymarginalis Schroeder, Reticulinella sp., Trochospira avnimelechi Hamaoui and Saint-Marc, Daxia cenomana Cuvillier and Szakall, Murgeina apula (Luperto Sinni), Nezzazata sp., Nezzaztinella picardi (Henson), Whiteinella baltica Douglas and Rankin and Heterohelix reussi (Chushman), Rotalipora appenninica (Renz), Rotalipora sp., Calcisphaerula sp. Planktons are investigated rarely in this biozone. Actually, benthic foraminifers are very abundant in Biozone 2. Many authors have assigned the mentioned faunal assemblage to the Middle to Upper Cenomanian [13,17,18]. According to the foraminiferal constituents of this biozone, its age is Late Cenomanian (Figure 8).
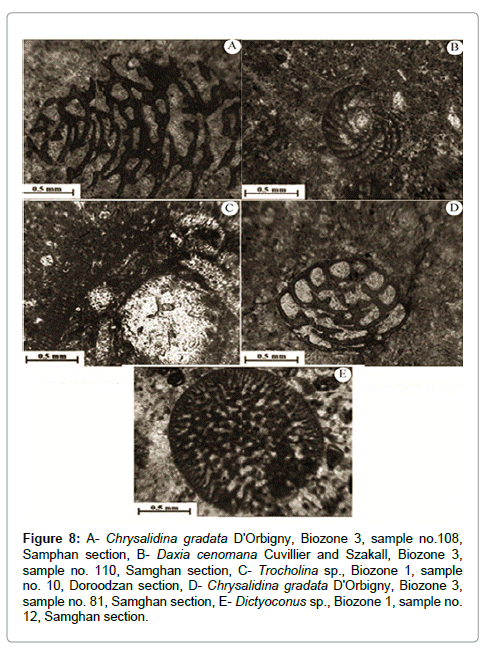
Figure 8: A- Chrysalidina gradata D'Orbigny, Biozone 3, sample no.108, Samphan section, B- Daxia cenomana Cuvillier and Szakall, Biozone 3, sample no. 110, Samghan section, C- Trocholina sp., Biozone 1, sample no. 10, Doroodzan section, D- Chrysalidina gradata D'Orbigny, Biozone 3, sample no. 81, Samghan section, E- Dictyoconus sp., Biozone 1, sample no. 12, Samghan section.
Doroodzan biostratigraphy
The lower lithostratigraphic limit of the Sarvak Formation in this section is conformable with the Kazhdumi Formation and overlies by thin-bedded limestone of Ilam Formation disconformably. The thickness of the Sarvak in the Doroodzan section is approximately 207 meters. It includes a succession of thin to medium-bedded grey limestone with rudist remains and Exogyra sp. Iron nodules are recognizable at the uppermost of the lithostratigraphic boundary.
Based on the microscopic investigations, some benthic and planktonic foraminifera have been identified at the level of species and genus. Benthic foraminifera include Orbitolina sp., Trocholina sp., Praealveolina cretacea (D´Archiac), Dicyclina schlumbergeri Munier-Chalmas, Nezzazata conica Smout, Nezzazata sp., Dictyoconus sp., Chrysalidina sp.; and planktonic foraminifera include Favusella washitensis (Carsey) , Ticinella roberti (Gandolfi), Calcisphaerula sp., Rotalipora sp. Based on the foraminiferal constituents, two assemblage biozones have been suggested for this stratigraphic section, which are Orbitolina sp. and the Nezzazata conica Assemblage Zone as Biozone 1, and the other one is named the Pseudolituonella reicheli and Nezzazata sp. Assemblage Zone (Biozone 2) (Figure 9).
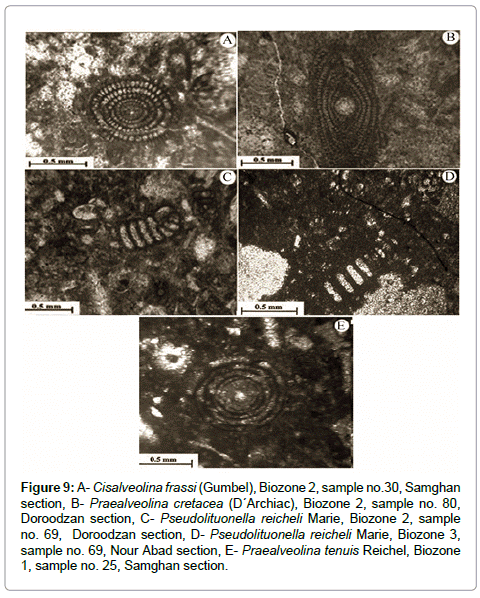
Figure 9: A- Cisalveolina frassi (Gumbel), Biozone 2, sample no.30, Samghan section, B- Praealveolina cretacea (D´Archiac), Biozone 2, sample no. 80, Doroodzan section, C- Pseudolituonella reicheli Marie, Biozone 2, sample no. 69, Doroodzan section, D- Pseudolituonella reicheli Marie, Biozone 3, sample no. 69, Nour Abad section, E- Praealveolina tenuis Reichel, Biozone 1, sample no. 25, Samghan section.
Biozone 1: (Orbitolina sp. and Nezzazata conica Assemblage zone)
The lithologic characteristics of this biozone consist of massive to thick-bedded grey limestone with iron nodules and thin-bedded dark gray limestone which extend about 130 meters in thickness. This biozone is described by the first appearance of Orbitoilina sp. and Nezzazata conica Smout. The associated foraminifers of this biozone are Orbitolina sp., Trocholina sp., Dicyclina schlumbergeri Munier- Chalmas, Dictyocunus sp., Chrysalidina sp., Pseudolituonella sp., Cuneolina sp. and Ticinella roberti (Gandolfi). The first occurrence of the mentioned foraminifers is synchronous with the last presence of Hemicylammina sigali Maync of the Kazhdumi Formation.
Aguilera-Franco [15] recognized Dicyclina schlumbergeri Munier- Chalmas from the Cenomanian strata of southwest Mexico. It should be noted that this taxon has a wide range which has been distinguished from the Campanian and Maastrichtian by many authors [1,49]. Nezzazata conica Smout was reported as a Lower to Middle Cenomanian taxon of Adriatic Platform succession (Croatia) by Husinec et al. [14] whereas it is recorded from the Morelos formation (SW of Mexico) in the Pseudorapydionina dubia zone (Early–Mid Cenomanian) by Aguilera-Franco [15]. Also, it has been documented by Mancinelli and Chiocchini [50] from the Lower–Upper Cenomanian boundary sequence of Monte Cairo (central–southern Italy) in association with Praealveolina tenuis Reichel. Veli? [18] has documented this species from the Early Cenomanian faunal assemblage of Karst Dinarides (SE Europe). Similar data has been received from northwest Syria [14,22,28] has distinguished this taxon with Nezzazata gyra Smouth from the southern part of the Mesozoic Adriatic Platform whereas Chiocchini et al. [51] recognized Nezzazata conica Smout from the Upper Cenomanian strata of Lazio meridionale (Italy). Other similar biostratigraphy studies have been recorded by other workers [28,38,50,51]. These biostratigraphic data reveal different stratigraphic ranges of Nezzazata conica Smout in the Tethyan realm. However, the faunal assemblage of the Adriatic platform is very close to the faunal characteristic value of the Zagros. Therefore, it confirms the Early–Mid Cenomanian age of this biozone (Figure 10).
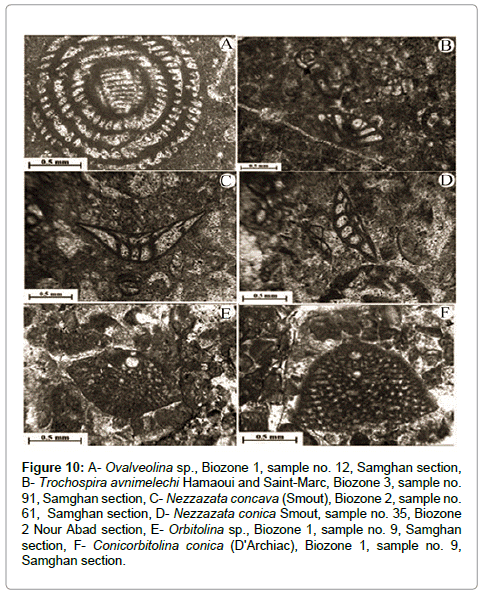
Figure 10: A- Ovalveolina sp., Biozone 1, sample no. 12, Samghan section, B- Trochospira avnimelechi Hamaoui and Saint-Marc, Biozone 3, sample no. 91, Samghan section, C- Nezzazata concava (Smout), Biozone 2, sample no. 61, Samghan section, D- Nezzazata conica Smout, sample no. 35, Biozone 2 Nour Abad section, E- Orbitolina sp., Biozone 1, sample no. 9, Samghan section, F- Conicorbitolina conica (D'Archiac), Biozone 1, sample no. 9, Samghan section.
Biozone2: (Pseudolituonella reicheli. and Nezzazata sp. Assemblage Zone)
Generally, this biozone is characterized by medium- to thickbedded dark grey limestone with pelecypod fragments and iron nodules. In addition, the diagnosed sublithofacies is very close to the Sarvak Formation of the Samghan section. Generally, bioclastic wackestone and Miliolid packstone are common sublithofacies of Biozone 2. Additionally, the mentioned lithofacies are common lithofacies of shallow water carbonate system in the Cenomanian strata of the Zagros region.
This biozone is marked by the first appearance of Pseudolituonella reicheli Marie and Nezzazata sp. Diagnosed foraminifers of this biozone are comprised of Rotalipora sp., Nezzazata sp., Dictyoconus sp., Cuneolina sp. and Praealveoloina cretacea (D´Archiac) have been recognized in this biozone. Actually, the mentioned foraminifers are distinguished in bioclast wackestone and packstone. Aguilera- Franco et al. [15] have documented Pseudolituonella reicheli from the Cenomanian sequence of the Morelos basin (southern Mexico) whereas Sari et al. [37] have reported this species from the Middleto Upper Cenomanian sediments of southwest Turkey. Futhermore, Husinec et al. [13] have documented Pseudolituonella reicheli as a Mid-Cenomanian foraminifer association (e.g. Nummoloculina regularis Philipson, Spiroloculina cretacea Auctt, Chrysalidina gradata D'Orbigny) in the upper part of Baldrin stratigraphic section island of Lošinj (Croatia). Borghi and Pignatti [41] have recorded this taxon from the uppermost Cenomanian succession of Apulia (southern Italy). Moreover, the study by Cadilac and Canerot [52] has recognized Praealveolina cretacea (D´Archiac) in the upper part of the Cenomanian sequence of Iberia and Catalonia (Spain). Cadillac and Canerot [52] reported Praealveolina cretacea (D´Archiac) from Upper Cenomanian sediments of Spain. Moreau [53] has recorded Praealveoloina cretacea (D´Archiac) from the Mid to Upper Cenomanian succession of north Aquitaine (France). His study reflected Dicyclina schlumbergeri Munier- Chalmas as an association in this region. Based on the foraminiferal constituents of this zone, the age of this biozone is suggested as Early Cenomanian (Figures 11 and 12).
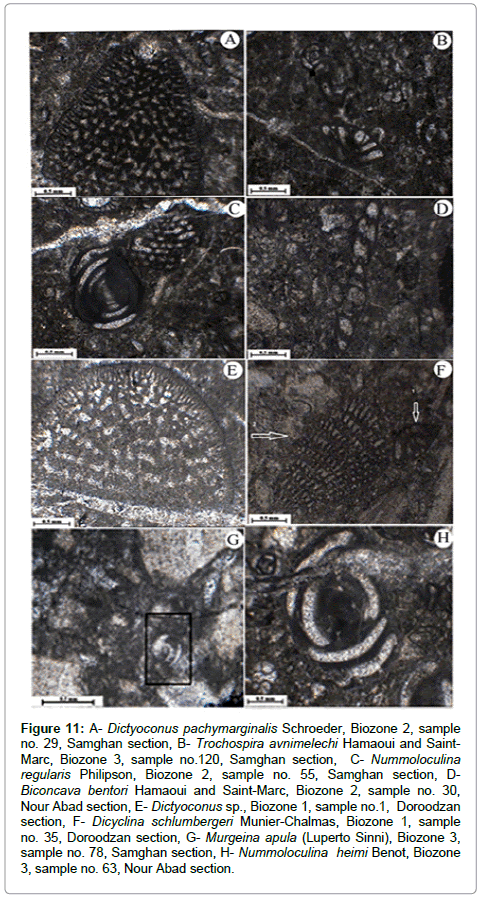
Figure 11: A- Dictyoconus pachymarginalis Schroeder, Biozone 2, sample no. 29, Samghan section, B- Trochospira avnimelechi Hamaoui and Saint- Marc, Biozone 3, sample no.120, Samghan section, C- Nummoloculina regularis Philipson, Biozone 2, sample no. 55, Samghan section, DBiconcava bentori Hamaoui and Saint-Marc, Biozone 2, sample no. 30, Nour Abad section, E- Dictyoconus sp., Biozone 1, sample no.1, Doroodzan section, F- Dicyclina schlumbergeri Munier-Chalmas, Biozone 1, sample no. 35, Doroodzan section, G- Murgeina apula (Luperto Sinni), Biozone 3, sample no. 78, Samghan section, H- Nummoloculina heimi Benot, Biozone 3, sample no. 63, Nour Abad section.
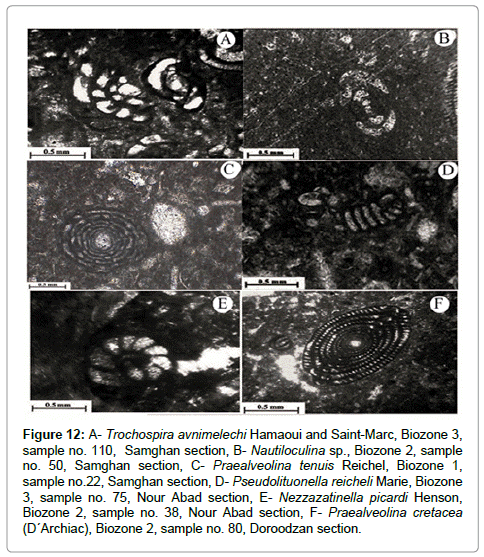
Figure 12: A- Trochospira avnimelechi Hamaoui and Saint-Marc, Biozone 3, sample no. 110, Samghan section, B- Nautiloculina sp., Biozone 2, sample no. 50, Samghan section, C- Praealveolina tenuis Reichel, Biozone 1, sample no.22, Samghan section, D- Pseudolituonella reicheli Marie, Biozone 3, sample no. 75, Nour Abad section, E- Nezzazatinella picardi Henson, Biozone 2, sample no. 38, Nour Abad section, F- Praealveolina cretacea (D´Archiac), Biozone 2, sample no. 80, Doroodzan section.
Results and Discussion
The vertical distribution of index foraminifers reveals various lower biostratigraphic limits in the studied stratigraphic sections. This study also confirms that the Sarvak Formation has established biozones that are different in their characteristic values. The upper biostratigraphic limit of the Sarvak is marked by an obvious disconformity. Unlike Wynd´s [7] suggestion that the reflected pelagic microfauna is expanded just in the Lurestan area, our record provides support that plankton is well developed in some of the studied stratigraphic sections (e.g. Arsenjan and upper part of Nour Abad stratigraphic sections). However, the biostratigraphic data of the four studied sections indicates that the Sarvak Formation’s established biozones consisted of benthic foraminifers. Actually, the determination of various lower biostratigraphic limits of the Sarvak Formation is an indication of the different sedimentary conditions of the studied sections along Fars platform. It should be noted that the well distributed planktons in the Arsenjan and Nour Abad sections are another proof confirming the paleoenvironmental divergence. Likewise, the first occurrence of planktons in the lower part of the Arsenjan Sarvak Formation reveals Upper Albian transgression in this area.
The biostratigraphic investigations of this work support well expanded Orbitolinid in the base portion of the some studied sections (e.g. the Nour Abad, Samghan, Doroodzan sections). Based on Vilas et al. [36], the distribution of Orbitolinids confirms the shallowest part of paleoenvironment of the mentioned sections, which is distinguished in the basal portion of the Sarvak Formation. Mancinelli and Chiocchini [50] referred Orbitolinids to back reef facies. Therefore, it is another proof of the domination of the shallow water carbonate system of Orbitolinid bearing strata of the Sarvak Formation in the studied sections. However, the age determination divergence of the Sarvak Formation in the studied sections is assigned to the paleomorphological aspect of the sedimentary basin and the tectonic effects which controlled the local transgressions and regressions in the Cenomanian times along the Fars platform. The occurrence of planktons in Arsenjan and Nour Abad is an indicator of the mentioned sea level fluctuations. Afghah and Shaabanpour Haghighi [54] have documented the fault control of biozonation in the Aptian carbonate sequence of the Nour Abad area. They explained the Kazeroon fault effect during sedimentation in Nour Abad region; eventually the Kazeroon fault had a tectonic effect similar to Aptian vertical displacement, whereas pelagic facies is recognized in whole Sarvak section of the Arsenjan region. The biostratigraphy of this section has not assisted the benthic assemblage fauna. Therefore, it is expected that there was transgression in the Arsenjan area during Mid-Albian through to Early Cenomanian age.
Conclusion
The difference between our new biostratigraphic data and previous works [4,7] is represented by the various biostratigraphic limits of the Sarvak Formation in the Fars platform. It should be noted that the vertical distribution of the Cenomanian foraminifer taxa is a function of the geologic setting of each stratigraphic section. The presence planktons of upper portion of the Nour Abad Sarvak Formation is an acceptable proof of vertical displacement of Kazeroon fault in the mentioned area during Late Cenomanian time. Therefore, it is approval geologic setting affected on stratigraphic distribution of upper Sarvak Formation foraminifers and other pelagic micrrofossils in Nour Abad section. Actually existence of planktons and other pelagic biofacies in studied area (e.g. Calcisphaerula innominata Benot) is an obvious biostratigraphic discrepancy with study of Wynd [7] which are determined in Arsenjan section. Biostratigraphic study on the mentioned section represents well distributed of pelagic facies during Late Albian through Early Cenomanian. Therefore contrary to previous works [7,8], pelagic facies is expanded in some regions of Fars platform. This study confirms similar characteristic value of benthic faunal assemblage in Nour Abad, Samghan and Doroodzan sections which represents close similarities with works of Kalantari [4]. It is necessary to note that well diversified benthic taxa are generally determined in Samghan section. The recognition of identified foraminifers supports well diversified Nezzaztids and Alveolinids remarkably in shallow water deposits of the Sarvak Formation. Orbitolinids are not valuable in the biostratigraphic aspect of the Cenomanian strata of studied sections. Additionally, they are distinguishable in Lower Cenomanian of Samghan and Doroodzan sections. The biozonation of the Sarvak Formation reveals the existence of obvious disconformity on upper biostratigraphic limit of Sarvak Formation. Although age dtermination of the Arsenjan Sarvak Formation section confirms Late Albian to Early Cenomanian, the lower biostratigraphic limit of the other stratigraphic of Sarvak Formation supports Early Cenomanian age. Therefore the lower biostratigraphic limit of the Sarvak Fomation is not synchronous in Fars platform exposures.
References
- Afghah M, Farhoudi G (2012) Boundary between Upper Cretaceous and Lower Paleocene in the Zagros Mountain Ranges of Southwestern Iran. ActaGeologicaSinica86: 325-338.
- Van Buchem FSP, Allan TL, Laursen GV, Lotfpour M, Moallemi A, et al. (2010) Regional stratigraphic architecture and reservoir types of the Oligo-Miocene deposits in the Dezful Embayment (Asmari and Pabdeh Formations) SW Iran. Geol Society, London, Special publication 329: 219-263.
- Tarbiyejou S, Afghah M, Sarnadi R, Yamini AR (2014) The determination and identification microfacies collection and sedimentary environments of Ilam and Sarvak reservoirs in Sarvestan oil field, proceeding of 32nd National and 1st International Geoscience Congress Petroleum Geology, Iran, Ahvaz 174.
- Kalantari A (1976) Microbiostratigraphy of Sarvestan area. National Iranian Oil Company, publication no. 5, Iran.
- James GA, Wynd JG (1965) Stratigraphic nomenclature of Iranian oil consortium agreement area. American Association of Petroleum Geologists Bulletin 49: 2182-2245.
- Ghavidel Syooki M, Motiei H, Akbar M, Rezaei A (2003) The petroleum geology of Iran. Reservoir Optimization Conference 1-31.
- Wynd JG (1965) Biofacies of the Iranian Oil Consortium Agreement Area. IOOC Report No.1082, (Unpublished).
- Motiei H (1992) Stratigraphy of Iran, Zagros stratigraphy. Geology and Mineral exploration Organization, Geological Survey of Iran 583.
- Afghah M, Ghiyasi AA (2013) Biostratigraphy of Gurpi Formation (Late Cretaceous) in Interior Fars Area, Central Zagros (Southwestern Iran). J Earth SciClim Change 4: 1-10.
- Tehrani KK, Fonooni B (1994) New Investigations in Microbiostratigraphy of Sarvak Formation in Fars & Khuzestan Region, Geological Survey of Iran. Geosci J 3: 2-15.
- Afghah M, Dehghanian MS (2007) Qualitative and quantitative study of the microfacies of the Sarvak Formation Kuh-e Khaneh-Kat and Kuh-e Shah-Neshin Sections of Fars Province. J Appl Geology 3: 7-16.
- Razin P, Taati F, van Buchem FSP (2010) Sequence stratigraphy of Cenomanian-Turonian carbonate platform margin (Sarvak Formation) in the High Zagros, SW Iran: an outcrop reference model for Arabian Plate. The Geological Society, London, Special publication 329: 187-218.
- Husinec A, Velic I, Fucek L, Vlahovic I, Maticec D, et al. (2000) Mid Cretaceous Orbitolinid (foraminiferida) record from the Islands of Cres and Losinj (Croatia) and its regional stratigraphic correlation. Cretaceous research 21: 155-171.
- Husinec A, Velic I, Sokac B (2009) Diversity patterns in mid-Cretaceous benthic foraminifers and dasyclad algae of the southern part of Mesozoic Adriatic Platform, Croatia. In: Demchuk, J. and Gary, A. (eds.) Geologic problem solving with microfossils, SEPM Special Publication 93: 153-170.
- Aguilera-Franco N (2003) Cenomanian-Coniasian zonation (foraminifers and calcareous algae) in the Guerrero-Morelos basin, Southern Mexico. Revista Mexicana de Ciencias Geological 20: 202-222.
- Premoli Silva I, Verga D (2004) Practical manual of cretaceous plankton icforaminifera, International school on planktonic foraminifera 283.
- Shahin A (2007) Oxygen and carbon isotopes and foraminiferal biostratigraphy of the Cenomanian-Turonian succession in GabalNezzazat, southwestern Sinai, Egypt. Revue de Paléobiologie 26:359-379.
- Velic I (2007) Stratigraphy and palaeobiogeography of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe). GeologicaCroatica60: 1-113.
- Chiocchini M (2008) New benthic foraminifers (Miliolacea and Soritacea) from the Cenomanian and Upper Turonian of the Monte Cairo (Southern Latium, Central Italy), Mem. Descr. Carta Geol 171-202.
- Gertsch B, Keller G, Adatte T, Berner Z, Kassab AS, et al. (2008) Cenomanian-Turonian transition in a shallow water sequence of the Sinai, Egypt, Springer. Int J Earth Sci 99: 165-182.
- Vincent B, Van Buchem FSP, Bulot LG, Immenhauser A, Caron M, et al. (2010) Carbon-isotope stratigraphy biostratigraphy and organic matter distribution in the Aptian-Lower Albian successions of southwest Iran (Dariyan and Kazhdumi formations). GeoArabia, Special publication 1: 139-197.
- Ghanem H, Mouty M, Kuss J (2012) Biostratigraphy and carbon-isotope stratigraphy of the uppermost Aptian to Upper Cenomanian strata of the South Palmyrides, Syria. GeoArabia17: 155-184.
- Orabi OH, Osman RA, El Qot GM, Afify AM (2012) Biostratigraphy and stepwise extinctions of the larger foraminifera during Cenomanian (Upper Cretaceous) of Gebel Um Horeiba (Mittla Pass), west-central Sinai. Egypt Revue de Paléobiologie 31: 303-312.
- Al-Dulaimy RT, Al-Sheikhly SS (2013) Biostratigraphy of Mishrif formation from well Amarah-1 southwestern Iraq. Iraqi Bull Geol Mining 9: 1-14.
- Alavi M (2004) Regional stratigraphy of the Zagros fold-thrust belt of Iran and its proforeland evolution. American Journal of Science 304: 1-20.
- Stoecklin J (1968) Structural history and tectonics of Iran: a review. American Association Petroleum Geologists bull 52: 1229-1258.
- Falcon NL (1974) SouthernIran: ZagrosMoutains, in Mesozoic-Cenozoicorogenicbelts: data fororogenicstudies: Alpine-HimalayanOrogens. Geological Society of London, Special Publications 4: 199-211.
- Ghanem H, Kuss J (2013) Stratigraphic control of the Aptian-Early Turonian sequences of the Levant Platform, Coastal Range, Northwest Syria. GeoArabia18: 85-132.
- Vaziri SH, Snowbari-Daryan B, Ghadimvand NK (2005) Lithofacies and microbiofacies of the Upper Cretaceous rocks (Sadr unit) of Nakhlak area in northeastern Nain, Central Iran. Journal of Geoscience, Osaka city University 48: 71-80.
- Omaña L, López Doncel R, Torres JR, Alencaster G (2012) Biostratigraphy and paleoenvironment of the Cenomanian/Turonian boundary interval based on foraminifera from W Valles-San Luis Potosà Platform, Mexico. Micropaleontol58: 457-485.
- Calonge A, Caus E, López-Carrilo D (2003) Biostratigraphy of the shallow-water Cenomanian deposits from the Iberian ranges (Spain). AAPG Search and Discovery Article #90017©2003 AAPG International Conference, Barcelona, Spain 21-24.
- Bosellini A, Morsilli M, Neri C (1999) Long-term event stratigraphy of the Apulia Platform margin (Upper Jurassic to Eocene, Gargano, Southern Italy). J SedimentRes69: 1241-1252.
- Sartorio D, Tunis G, Venturini S (1992) Open circulation facies in the Cenomanian of the northeastern margin of the Friuli Platform: the Ludrio Valley case (NE Italy). GeologicaCroatica45: 87-93.
- Simmons MD, Whittaker JE, Jones RW (2000) Orbitolinids from Cretaceous sediments of the Middle East - a revision of the F.R.S. Henson and Associates Collection. In: Hart MB, Kaminski MA, Smart CW (edn) Proceedings of the Fifth International Workshop on Agglutinated Foraminifera. Grzybowski Foundation Special Publication 7: 411-437.
- Tertis L, Buccur II, Sasaran E, Ghergari L (2002) New remarks on the lithology and biostratigraphy of the Cretaceous deposits from Istria Depression (Black Sea shelf). StudiaUniversitatis Babes-BolyaiGeologia 75-84.
- Vilas L, Masse JP, Arias C (1995) Orbitolina episodes in carbonate platform evolution: the early Aptian model from SE Spain. Palaeogeogr Palaeocl Palaeoecol 119: 35-45.
- Sari B, Tasli K, Özer S (2009) Benthonic foraminiferal biostratigraphy of the Upper Cretaceous (Middle Cenomanian-Coniacian) sequences of the BeyDaglari carbonate platform, Western Taurides, Turkey. Turkish J Earth Sci 18: 393-425.
- Velic I, Vlahovic I (1994) Foraminiferal assemblages in the Cenomanian of the Buzet-Savudrija Area (Northwestern Istria, Croatia). Geologica Croatica 47: 25-43.
- Mouty M, Al-Maleh A K, Laban HA (2003) The Middle Cretaceous in the Palmyrides chain. Geodiversitas 25: 429-443.
- Hernández-Romano U, Aguilera-Franco N, MartÃnez-Medrano M, Barceló-Duarte J (1997) Guerrero-Morelos Platform drowning at the Cenomanian-Turonian boundary, Huitziltepec area, Guerrero State, southern Mexico. Cretaceous Research 18: 661-686.
- Borghi M, Pignatti JS (2006) Taberinabingistani Henson, 1948 (Foraminifera) from the Upper Cenomanian of the Apulia (southern Italy): a new record. GeologicaRomana39: 89-94.
- Filkorn HF, Scott RW (2011) Microfossils, paleoenvironments and biostratigraphy of the Mal Paso Formation (Cretaceous, upper Albian), State of Guerrero, Mexico. Revista Mexicana de Ciencias Geológicas 28: 175-191.
- Keller G, Pardo A (2004) Age and paleoenvironment of the Cenomanian-Turonian global stratotype section and point at Pueblo, Colorado. Mar Micropaleontol51: 95-128.
- Cetean CG, Balc R, Kaminiski MA, Filipscu S (2008) Biostratigraphy of the Cenomanian-Turonian boundary in the Eastern Carpathians (Dâmbovita Valley): preliminary observations. StudiaUniversitatisBabes-Bolyai, Geologia 53: 11-23.
- Fucek L, Velic I, Vlahovic I, Oštric N, Korolija B ,et al. (1995) Novipodaci o stratigrafijidonjekredeotokaCresa. In: Vlahovic´ I, Velic´ I, Syparica M (edn) HrvatskiGeolos?kiKongres, Opatija 1995, Zbornikradova 1 [The First Croatian Geological Congress, Opatija 1995, Proceedings 1] 167-172.
- Berthou PY (1984) Albian-Turonian stage boundaries and subdivisions in the western Portuguese Basin, with special emphasis on the Cenomanian-Turonian boundary in the ammonite facies and rudistfacies. Bull GeolSoc Denmark 33: 41-55.
- Calonge A, Caus E (1996) Particularidades de los macroforaminÃferos del Cenomaniense de la Cardillera Ibérica: implicaciones paleobiogeográcas. Geogaceta 20: 191-194.
- Hart MB, Callapez PM, Fisher JK, Hannant K, Monteiro JF, et al. (2005) Micropalaeontology and stratigraphy of the Cenomanian/Turonian boundary in the Lusitanian Basin, Portugal. J Iberian Geol 31: 311-326.
- Permoli Silva I, Nicora A, Vanneau AA (1995) Upper Cretaceous larger foraminifer biostratigraphy from WodejebatoGuyot, site 873 through 877. Proceedings of the Ocean Drilling Program, Scientific Results 144: 171-198.
- Mancinelli A, Chiocchini M (2006) Cretaceous benthic foraminifers and calcareous algae from Monte Cairo (southern Latium, Italy),Bollettinodella Società Paleontologica Italiana45: 91-113.
- Chiocchini M, Coccia B, Mancinelli A, Romano A, Urgera A (2004) Microbiostratigrafiaedevoluzionepaleogeograficadel Mesozoico del Monte Cairo e di Vallerotonda(Laziomeridionale, Italia). Studi Geol Camerti, NuovaSerie 2: 1-18.
- Cadillac H, Canerot J (1978) Données stratigraphiques et paléogéographiques nouvelles sur leCretacé supérieur aux confins des chaines Ibérique et Catalane (Espagne). Acta Geologica Hispanica 13: 155-158.
- Moreau P (1996) Analysis of the 4 Cenomaniantransgression over the north-western border of the Aquitaine basin. Géologie de la France 1: 3-16.
- Afghah M, Haghighi SA (2014) Aptian biostratigraphy in south Zagros Basin, southwest Iran. Elsevier, Geoscience Frontiers5: 277-288.
Citation: Afghah M, Yousefzadeh A, Shirdel S (2014) Biostratigraphic Revision of Middle Cretaceous Succession in South Zagros Basin (SW of Iran). J Earth Sci Clim Change 5: 216. DOI: 10.4172/2157-7617.1000216
Copyright: ©2014 Afghah M, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Select your language of interest to view the total content in your interested language
Share This Article
Recommended Journals
Open Access Journals
Article Tools
Article Usage
- Total views: 17881
- [From(publication date): 10-2014 - Aug 24, 2025]
- Breakdown by view type
- HTML page views: 12966
- PDF downloads: 4915