Taphonomy of Aptian-Albian Beds in the Gebel Mistan, Maghara Area, Northern Sinai, Egypt
Received: 28-Feb-2013 / Accepted Date: 15-Mar-2013 / Published Date: 18-Mar-2013 DOI: 10.4172/2157-7617.1000135
Abstract
We can discover much information and many of the secrets of life in the past through studying fossils. Taphonomy can currently be used to reveal this information, but unfortunately, this branch of paleontology has not yet been wellexploited in Egypt, with only a small number of studies having been conducted. The present study is an attempt to understand the taphonomic conditions under which several Aptian-Albian beds from the Gebel Mistan region were deposited. The order of the beds from the base to the top is as follows; 1- Nerinea bed (Upper Aptian), 2- Coral bed (Upper Aptian), 3-Acteonella bed (Middle Albian) and Nerinea bed (Middle Albian). For each bed, the lithology, thickness, faunal content, state of preservation, packing density, orientation patterns, fragmentation, bioerosion and encrustation were described. The present study led to the conclusion that the four beds (Upper Aptian- Middle Albian) were deposited in a shallow, slightly–to-very-worm-marine environment with clear to agitated water.
Keywords: Lower cretaceous; Nerinea; Corals; Acteonella
9540Introduction
The Lower Cretaceous successions, which have outcrops along the northern flanks of the Gebel Maghara Dome, exhibit a rich and diversified assemblage of macrofossils [1]. The Lower Cretaceous marine sequences are best exposed in the Maghara area [1,2]. Although the Lower Cretaceous fauna of the northern Sinai is of high quality [3], the taphonomic attributes of these fossils have received little attention [1,4].
Taphonomic studies that have previously been conducted in Egypt include those of [1,4-10]. No taphonomic studies have been carried out on the Lower Cretaceous macrofauna, except by [1], who studied three different phenomena (inquilinism, predation and taphonomy) in certain cephalopod shells collected from the Aptian-Albian sections of Gebel Lagma and Gebel Manzour in the Maghara region.
The goal of this paper is to study the taphonomic conditions of sclerobionts in the four Lower Cretaceous beds from Gebel Mistan to understand and interpret the paleobiology and paleoenvironment of these beds.
Stratigraphic setting
The northern Sinai is characterized by the presence of excellent outcrops. These outcrops have allowed for the implementation of significant research, especially concerning stratigraphy and sedimentary structure. The exposed Lower Cretaceous rocks of the northern Sinai around Gebel Maghara have received considerable attention from the geologists, beginning with [11] and continuing with [12-38].
In Gebel Mistan, the Lower Cretaceous succession is subdivided into two lithostratigraphic units, namely, the Risan Aneiza and the Halal formations, as further described below from base to top (Figure 1).
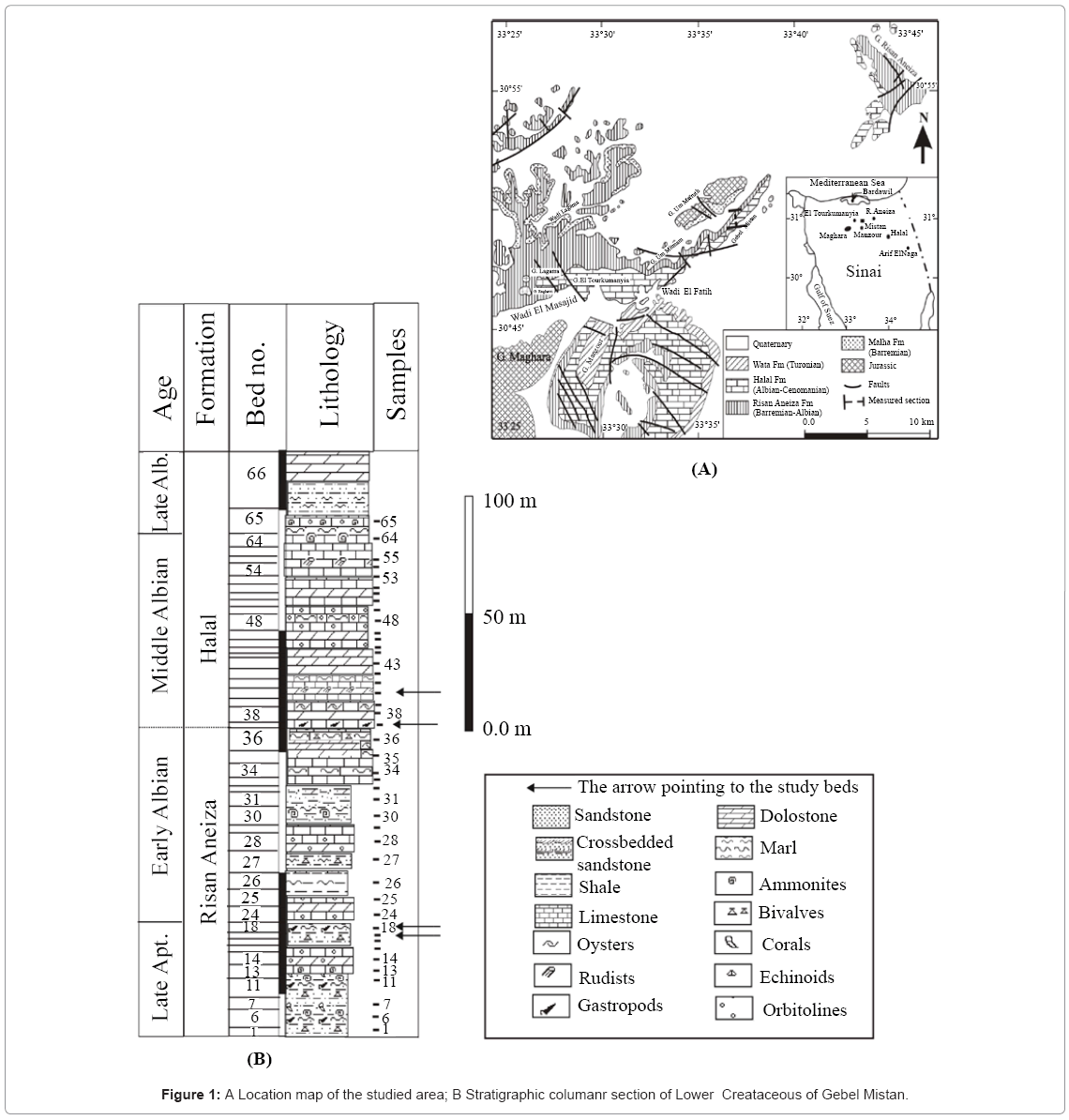
Figure 1: A Location map of the studied area; B Stratigraphic columanr section of Lower Creataceous of Gebel Mistan.
Risan Aneiza Formation: Underlies the Halal Formation. This formation’s base is unexposed, and it consists mainly of green shale, limestone and marl with sandstone interbeds with a thickness of 160 m.
Halal Formation: Consists predominantly of white and grey dolostones that are commonly intercalated with thin marl and have limestone interbeds. This formation conformably overlies the Risan Aneiza Formation. The Halal Formation has a thickness of 140 m.
Materials and Methods
The Aptian-Albian sediments in Gebel Mistan are highly fossiliferous, being composed of invertebrate macrofossils, predominantly bivalves and gastropods. Most of the fauna was wellpreserved especially what has been collected from the marl beds and has been included in the present research. The author selected four beds (two from the Upper Aptian and two from the Middle Albian) for an extensive taphonomic analysis. In the field, for each bed, the lithology, thickness, faunal content, state of preservation, packing density, orientation patterns, fragmentation state, bioerosion level and encrustation status were described. The material studied is housed at the Geology Department, Suez Canal University, Ismailia, Egypt.
Results
The Upper Aptian Nerinea bed
The bed ranges in thickness from 0.5 to 1.5 m and consists of a hard, yellowish brown limestone. The faunal content of this bed is composed mainly of complete, well-preserved, small, lensoid shaped, densely packed, randomly oriented, non-bioeroded, non-encrusted shells of Nerinea mistanensis Awad 1952 and Nerinea maghrensis Abbass 1963 in addition to the shells of other gastropods, such as Tylostoma (Tylostoma) magharensis Abbass 1963, Pyrazus (Echinobethra) magharensis Abbass 1963, Pyrazus (Echinobethra) sexangulatus Zék. Permut1852, Pseudomesalia quadrilineata Mahmoud 1956 and Turritella (Torquesia) moradi Abbass 1963. The associated bivalves are Ceratostreon flabellatum (Goldfuss 1833), Nucula (Nucula) margitifera Douvillé 1961 and Maghrebella deserti (Douvillé 1961). In general, the associated gastropod and bivalve fauna is poorly sorted, moderately fragmented, loosely packed and articulated, and certain gastropods show traces of bioerosion, while smaller bivalves show evidence of encrustation. Most of the associated gastropods and bivalves are detected as internal molds (Plate 1A).

Plate 1: A-D Field photographs of the study beds A Upper Aptian Nerinea bed; B Upper Aptian coral bed; C Middle Albian Acteonella bed; D Middle Albian Nerinea bed.
The lower (5 cm) portion of this bed consists of clusters of Chondrodonta sp. Their shells are randomly oriented and are large, highly fragmented and disarticulated, exhibiting no evidence of either bioerosion or encrustation.
The Upper Aptian coral bed
This bed is consists of a yellowish brown sandy limestone and measures 50 cm in thickness. The bed overlies the Nerinea bed and is composed principally of the corals Epistreptophyllum manzourensis Gameil and Aly 2004, Epistreptophyllum sp., Trochosmilia cretacea Gameil and Aly 2000, Mixastraea sp., Paracycloseris sp. and Montlivaltia sp. The coral fragments are loosely packed, relatively highly fragmented, randomly oriented and well-preserved (Plate 1B).
Most of the associated macrofauna collected are internal molds and are articulated, moderately fragmented, patterned in a random orientation, moderately preserved and poorly sorted. The echinoids and certain of the gastropods show traces of bioerosion, while a portion of the oyster shells are encrusted with small organisms.
The Middle Albian Acteonella bed
This bed consists primarily of a yellowish brown, dolomitic sandy limestone with a thickness of 1 m. The bed represents a monotaxic accumulation of the Acteonella delgadoi Choffat 1901 (Plate 1C). The Acteonella shells are small-to medium-sized, densely packed, moderately sorted, randomly oriented and relatively fragmented and exhibit no evidence of bioerosion or encrustation.
The Middle Albian Nerinea bed
The lithology of this bed is a white, hard dolomitic limestone approximately 75 cm in thickness. The bed is a monotaxic collection of the Nerinea abbassi Abbass 1963 shadwished between two rudist beds. The Nerinea shells are unimodal, perpendicular to the bed in the section form, densely packed, relatively fragmented, well-sorted and well-preserved and they are neither bioeroded nor encrusted (Plate 1D).
Interpretation
According to the field and taphonomic observations of the fossils (Table 1) contained in the four beds studied, their historical accumulation can be interpreted as follows:
| Studied beds | Nerinea bed | Corals bed | Acteonella bed | Nerinea bed |
| Properties | ||||
| 1- Age | Upper Aptian | Upper Aptian | Middle Albian | Middle Albian |
| 2- Bed no. | 17 | 18 | 37 | 40 |
| 3- Thickness | 0.5-1.5 m | 50 cm | 1 m | 75 cm |
| 4- Lithology | Yellowish brown, hard limestone | Yellowish brown, sandy limestone | Yellowish brown, dolomitic sandy limestone | White, hard dolomitic limestone |
| 5- Assemblage | Polytaxic | Polytaxic | Monotaxic | Monotaxic |
| 6- State of preservation | Well | Well | Moderate | Well |
| 7- Orientation pattern | Random | Random | Random | unimodel |
| 8- Fragmentation | Moderate | Relatively high | Low | Low |
| 9- Packing | Dense | Loose | Dense | Dense |
| 10- Sorting | Poor-moderate | Poor | Moderate | Good |
| 11- Geometry | Lens | Bed | Bed | Bed |
Table 1: The main field and taphonomic properties of the study beds.
The Upper Aptian Nerinea bed
The high quality and complete preservation of the Nerinea shells, their dense packing, random orientation and absence of bioerosion and encrustation might have been due to a short post-mortem period, a rapid burial rate after their death, lack of transportation, a relatively low sedimentation rate and-preservation in situ.
With respect to the associated fauna, their taphonomic signatures suggest that their preservation time and location were not the same as those of Nerinea shells. Perhaps the associated fauna were deposited after the Nerinea shells, or they were transported from another place, although not from too great a distance. The shells indicate a long postmortem period.
The random orientation, high fragmentation, high disarticulation and absence of bioerosion and encrustation of the narrower chondrodonta bed (5 cm) indicate a repetitive reworking in situ, a short residence time on the sea floor, rapid burial and a rapid rate of sedimentation.
The Upper Aptian coral bed
The taphonomic features of the coral fragments, such as high fragmentation, loose packing, good preservation and evidence of bioerosion and encrustation on certain of the coral skeletons? associated macrofauna might have resulted from a slow burial rate, a relatively long post-mortem period and transport over only short distances.
The Middle Albian Acteonella bed
The dense packing of the Acteonella shells, their moderate sorting, random orientation, low fragmentation and the absence of bioerosion and encrustation are strong evidence of the preservation of the shells in situ, a short post-mortem period, a rapid burial rate and their having been reworked more than once in situ with no evidence of transport.
The Middle Albian Nerinea bed
The manner of accumulation of the Nerinea shells with evidence of dense packing, low fragmentation and good preservation might indicate a relatively short distance of transportation by strong currents, a slow rate of sedimentation and a slow burial rate after death. Additionally, the absence of bioerosion and encrustation are strong evidence of a short post-mortem period.
Inferred depositional environment of the study area
Many paleontologists are interested in studying the depositional environment of the Lower Cretaceous in the northern Sinai in Egypt. The best-known studies are those of [33,39-43].
For the current study, the depositional environment of the Lower Cretaceous rocks has been examined for the first time in Egypt, and this environment has been illuminated by the study of the taphonomic characteristics of the fossils. Through the examination and evaluation of the four beds representing the Upper Aptian to Middle Albian in Gebel Mistan, it was determined that the Upper Aptian Nerinea and coral beds were deposited under relatively high sedimentation rates at a water depth of no more than 15 m in a notably warm climate (marked by the presence of corals and diversity of macrofauna) lacking large predators (scarcity or absence of bioerosion and traces of predation) during organisms´ lives.
As for the Middle Albian Acteonella and Nerinea beds, their depositional environment appears to have featured a low sedimentation rate in water depth deeper than that found in the depositional environment of the Upper Aptian beds (presence of dolomitic limestone) with slightly warm and slightly turbulent water (lack of diverse macrofauna) that also lacked large predators (absence of bioerosion and predation traces) during organisms´ lives.
Acknowledgements
Many thanks to everyone in Journal of Earth Science & Climatic Change who has contributed toward the progress of this work especially Gracia S. Oliver, Assistant Managing Editor to respond to any request I asked her. I am deeply grateful to Rituparna Bose, Editor, Adjunct Assistant Professor, City University of New York, USA for critical review of the manuscript. Deep and grateful thanks to the reviewers for their critical review of the manuscript and useful comments.
References
- EL-Shazly SH (2006) Inquilinism, predation and taphonomy of some Lower Cretaceous ammonites, North Sinai, Egypt. Egyptian journal of Paleontology 6: 159-191.
- Aly MF (2006) Aptian cephalopods from Gebel Abu Ruqum, North Sinai, Egypt. Egyptian journal of Paleontology 6: 89-123.
- Mekawy MS (2012) Unusual factor affecting the preservation of fossils from Northern Sinai, Egypt. Journal of Earth Science & Climate Change 3: 121.
- Mekawy MS (2007) Taphonomy of Cenomanian oysters from Gebel Yelleg, North Sinai, Egypt. Egyptian journal of Paleontology 7: 335-348.
- Kassab AS (1995) Biostratinomy of the Upper Cenomanian oyster storm shell beds from the Northeastern Desert of Egypt. Bollettino della Societá Paleontologica Italiana 43: 139-149.
- Abdel Hamid MAM (1999) Parasitism, abnormal growth and predation on Cretaceous echinoids from Egypt. Revue de Paléobiologie 18: 69-83.
- EL-Hedeny MM (2005) Taphonomy and paleoecology of the Middle Miocene oysters from Wadi Sudr, Gulf of Suez, Egypt. Revue de Paléobiologie 24: 719-733.
- EL-Hedeny MM (2007) Encrustation and bioerosion on Middle Miocene bivalve shells and echinoid skeletons: paleonvironmental implications. Revue de Paléobiologie 26: 381-389.
- EL-Hedeny MM, EL-Sabbagh AM (2007) Macro-borings on Late Cretaceous oysters of Egypt. Neues Jahrbuch für Geologie und Paläontologie 244: 273-286.
- EL Qot GM, Abdel-Gawad GI, Mekawy MS (2009) Taphonomy of Middle Jurassic (Bathonian) shell concentrations from Ras El Abd, west Gulf of Suez, Egypt. Journal of African Earth Sciences 54: 31-36.
- Douvillé MH (1916) Les terrains secondaires dans le massif du Moghara, a l´Est de l´lsthme de Suez. Mém. Acad. Sci., de l´lnstitute de France, Ser 2: 1-184.
- Moon FW, Sadek H (1921) Topography and geology of north Sinai, Egypt. Petroleum Research Bulletin (Cairo) 10: 1-154.
- Farag IAM (1947) Preliminary notes on the geology of Risan Aneiza. Bull Fac SciCairo Univ 26: 1-38.
- Awad GH (1952) The Nerineas of Sinai (with a note on the mode of these extinct gastropods). Bulletin de I´Institut du Désert d Egypte 2: 23-39.
- Moret L, Mahmoud IG (1953) Nouvelles observations stratigraphique et paleontologique sur I´Albien du Massif du Moghara (Sinai, Eygpte). Trav Lab Geol Univ Grenoble T 31: 267-272.
- Mahmoud GE (1955) Etudes paléontologiques sur la faune Crétacique du massif du Moghara (Sinai, Egypte). Publ. Desert Instd Egypte 8: 1-195.
- Said R, Barakat MG (1957) Lower Cretaceous foraminifera from Khashm El Mistan, Northern Sinai, Egypt. Micropaleontology 3: 39-47.
- Abbass HL (1962) A monograph on the Egyptian Cretaceous Cretaceous pelecypods. Geol Surv Miner Res Dept Palaeont Ser Mongr 1: 224
- Abbass HL (1963) A monograph on the Egyptian Cretaceous gastropods: Geological Survey and Mineral Research Department of the United Arab Republic, Palaeontological Series, Monograph 2: 146
- Ismail MM, Mansour AIY (1969) A microfacies study of the Lower Cretaceous of Gebel El Maaza, Northen Sinai. U.A.R. Alexandria Univ Fac Sci Bull 9: 19-32.
- Naim NE, Mansour AIY (1969) Ontogenetic and phyllogenetic studies on the genus Pseudomesalia. Douvillé Fac Sci Alex Univ 9: 555-565.
- Hegab OA, Hamama HH, Atia NA (1989) Stratigraphy, facies and environment of the Lower Cretaceous of Gebel Um Mitmam, Maghara area, North Sinai. Proc 2nd Conf Geol Sinai Development, Ismailia 110-120.
- Aboul Ela NM, Abdel-Gawad GI, Aly MF (1991) Albian fauna of Gabal, Maghara area, north Sinai, Egypt. Journal of African Earth Sciences 13: 201-220.
- Hassan MM, Abdel Hafez NA, Dardir A, Arian MA (1992) Geologic studies of the Cretaceous sedimentary rocks in Risan Aneiza-Gabal Amrar area, Northen Sinai, Egypt. Second Meeting of the Geology of Arab World, Cairo Univ. 353-364.
- Hamama H (1992) Aptian and Albian corals (Scleractinia) of the Northern Sinai, Egypt. Bull. Faculty of Science, Mansoura University 19: 87-113.
- Hamama H (1993) Albian ammonites of Northen Sinai, Egypt. Mans Sci Bull C (Natural Sci.) 20: 261-301.
- Hamza FH, Ismail AA, EL-Saadany MA (1994) Biostratigraphy and paleoecology of some Cretaceous exposures in Northen Sinai, Egypt.In EGPC 12th Petroleum Exploration and Production Conference, Cairo, 12-15 November 1994 482-495.
- Hewaidy AA, Azab MM, Kamel DE (1998) Macrobiostratigraphy of the Lower/Middle Cretaceous sequences in some parts of northen Egypt. Proceed 11th Symp Phaner Develop Egypt 39-77.
- Shahin A (2000) Albian ostracodes from Northen Sinai, Egypt. In 2nd Int. Conf. for Basic Sci. and Adv. Tec. (BAST-II), Assiut Univ., Assiut 129-156.
- Shahin A (2001) Contribution to the knowledge of the Early Cretaceous foraminiferal assemblages, their paleoecology and paleogeography in the Northen Sinai, Egypt. N Jb Geol Palaont Mh 221: 397-440.
- Hewaidy AA, Morsi AM (2001) Lower Cretaceous (Aptian-Albian) foraminifera and ostracoda from Northern Sinai, Egypt. Egyptian Journal of Paleontology 1: 229-252.
- Aly MF, Abdel-Gawad GI (2001) Early Cretaceous ammonites of Gebel Lagama, North Sinai, Egypt. Palaeontographica Abt A 262: 25-52.
- Bachmann M, Bassiouni MAA, Kuss J (2003) Timing of mid-Cretaceous carbonate platform depositional cycles, northern Sinai, Egypt. Palaeogeography, Palaeoclimatology, Palaeoecology 200: 131-162.
- Aly MF, Abdel-Gawad GI, Gabir MA (2005) Upper most Albian-Cenomanian ammonites from north Sinai, Egypt. Egyptian journal of Paleontology 5: 347-385.
- Abu-Zeid RH (2006) Biostratigraphic framework of some Lower Cretaceous outcrops from Northern Sinai, Egypt, based on cephalopods. Egyptian journal of Paleontology 6: 125-158.
- Mekawy MS, Abu-Zeid RH (2008) Lower Cretaceous molluscan fauna from North Sinai, Maghara area, Egypt. Egyptian journal of Paleontology 8: 291-334.
- Hamama H (2010) Barremian and Aptian Mollusca of Gabal Mistan and Gabal Um Mitmami, Al-Maghara area, Northern Sinai, Egypt. Journal of American Sciences 6: 1702-1714.
- EL-Shazly SH (2012) Macrofaunal and the trace fossil studies of the Lower Cretaceous rocks at Gebel Mistan, North Sinai, Egypt. Egyptian Journal of Geology 56: 289-332.
- Lewy Z (1990) Transgressions, regressions and relative sea level changes on the Cretaceous shelf of Israel and adjacent countries: A critical evaluation of Cretaceous global sea level correlations.Paleoceanography 5: 619-637.
- Bachmann M, Kuss J (1998) The Middle Cretaceous carbonate ramp of the northen Sinai: Sequence stratigraphy and facies distribution. In: Wright VP, Burchette TP (Eds.), Carbonate Ramps Geol Soc Lond Spec Publ 149: 253-280.
- Kuss J (1992) The Aptian-Paleocene shelf carbonates of Northeast Egypt and Southern Jordan: establishment and break-up of carbonate platforms along the Southern Tethyan shores.Z dt geol Ges 143: 107-132.
- Bachmann M, Bandel K, Kuss J, Willems H (1996) Sedimentary processes and intertethyal comparisons of two Early/Late Cretaceous ramp systems (NE-Africa and Spain). In: Reitner, J., Neuweiler, F., Gunkel, F. (Eds.), Global and Regional Controls on Biogenic Sedimentation, II. Cretaceous Sedimentation.Göttinger Arb. Geol. Paläont. Sb3,151-163.
- Kuss J, Bachnann M (1996) Cretaceous paleogeography of the Sinai peninsula and neighbouring areas.CR Acad Sci Paris 322: 915-933.
Citation: Mekawy MS (2013) Taphonomy of Aptian-Albian Beds in the Gebel Mistan, Maghara Area, Northern Sinai, Egypt. J Earth Sci Climate Change 4: 135. DOI: 10.4172/2157-7617.1000135
Copyright: ©2013 Mekawy MS. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Select your language of interest to view the total content in your interested language
Share This Article
Recommended Journals
Open Access Journals
Article Tools
Article Usage
- Total views: 15748
- [From(publication date): 5-2013 - Oct 09, 2025]
- Breakdown by view type
- HTML page views: 10820
- PDF downloads: 4928