Research Article Open Access
Frugivory in Bromelia Balansae (Bromeliaceae): The Effect of Seed Passage through the Digestive System of Potential Seed Dispersers on Germination in an Atlantic Rainforest, Brazil
Hipólito Ferreira Paulino-Neto*, Eduardo Nakano-Oliveira, Marcia Maria de Assis Jardim and João Vasconcellos-Neto
Professor of Biological Sciences and head researcher at the Laboratory of Ecology of pollination, Evolution and Conservation (LEPEC), University of the State of Minas Gerais (UEMG), Brazil
- *Corresponding Author:
- Paulino-Neto HF
Professor of Biological Sciences and head researcher at the Laboratory of Ecology of pollination
Evolution and Conservation (LEPEC)
University of the State of Minas Gerais (UEMG), Brazil
Tel: (035) 35296058
E-mail: hipolitopaulino@gmail.com
Received Date: December 22, 2016; Accepted Date: December 26, 2016; Published Date: December 30, 2016
Citation: Paulino-Neto HF, Nakano-Oliveira E, de Assis Jardim MM, Vasconcellos-Neto J (2016) Frugivory in Bromelia Balansae (Bromeliaceae): The Effect of Seed Passage through the Digestive System of Potential Seed Dispersers on Germination in an Atlantic Rainforest, Brazil. J Ecosys Ecograph 6: 224. doi:10.4172/2157-7625.1000224
Copyright: © 2016 Paulino-Neto HF, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Visit for more related articles at Journal of Ecosystem & Ecography
Abstract
Frugivory in Bromelia balansae (Bromeliaceae): the effect of seed passage through the digestive system of potential seed dispersers on germination in an Atlantic Rainforest, Brazil.
The Serra do Japi-SP Ecological Reserve is one of the last large areas of continuous forest in the interior of state São Paulo, and currently suffers great anthropic pressure due to the urbanization process in its surroundings. The dispersion of seeds by frugivorous mammals has great importance in the regeneration of altered areas and the composition of seed banks. In this work we identified some frugivores of B. balansae and tested the viability of seeds present in their feces. To identify the frugivores were installed "foot traps" near plants with mature fruits. Mammalian feces were collected and analyzed in order to find seeds of B. balansae. Three species of frugivores were identified: Cerdocyon thous, Nasua nasua and Didelphis sp. In captivity, mature fruits of B. balansae were offered to crab-eating foxes and coati to obtain seeds passed through the digestive tract of these animals. The seeds contained in the feces of these mammals were collected for germination experiments in the laboratory. As a control, seeds were extracted directly from mature fruits for germination tests. As main results, we verified that the seeds remained viable and intact after passing through the digestive tract of these two species. Seeds that passed through the digestive tract of N. nasua and C. thous germinated faster than seeds from fruits. Coati gut passage showed significantly higher germinability than both groups of seeds, without pulp and with pulp. But the germinability of seeds with pulp removal was significantly higher than seeds with pulp. However, seeds that were intact after crabeating foxes gut passage showed germination significantly higher than that seed with pulp, but showed no significant differences when compared to the seeds with removed pulp. Also, our data indicate that the coati gut passage has a higher positive effect on the seed germination of B. balansae than seed from foxes feces and mature fruits treatments. The plant architecture and leaf morphology of B. balansae restricts small size dispersers and favors the access of larger animals to the fruits, such as coatis and crab-eating foxes, ensuring the dispersion of seeds over long distances.
Keywords
Cerdocyon thous ; Didelphis sp.; Dispersal syndrome; Nasua nasua ; Plant architecture; seed removal; Seed germination; Seed dispersal; Terrestrial bromeliad
Introduction
Seed dispersal by vertebrates is fundamental for the reproductive mechanisms of many species of tropical plants [1]. The means of dissemination by birds and mammals are often associated with a plant’s investment in the nutritional quality of its fruit, as well as its color, shape and smell [2-4]. This strategy enhances the reproductive success of these plants by ensuring the dispersal of their seeds by animals [5-7]. Effective dispersal therefore depends on the amount of seeds consumed, on the probability of the seeds surviving the passage through the digestive tract and on the conditions for the plant become established as well as the phenology or rhythm of fructification [8,9]. The ingestion a fruit by an animal can have a positive or negative effect on the seeds. Thus, the removal of the pulp from the fruit, can remove possible inhibitions of germination and reduce the susceptibility to attack by fungi [10], thereby enhancing the lifetime and success of germination of the seeds. Alternatively, some or all of the ingested seeds may be destroyed during digestion [11] or can become unviable and hence be unable to contribute to the dispersion of the species [12,13]. The rate of germination could also be affected in different way depend on the animal dispersers and the time through gut passage [14,15]. Hence, depending on the animal species and on the fruits consumed, the passage through the digestive tract can reduce [16], increase the rate of seed germination, or leave it unaltered [13].
The dispersal of fruit is influenced by the behaviour, phyisiology and morphology of the frugivores [17]. The manner in which the fruit is handled and processed, and how long it remains in the gut all affect the viability and quantity of seeds that are transported [18]. The destiny of the seeds and their germination are affected both by the area in which the frugivores live and by their patterns of movement [18]. Thus, seeds may be moved far from the mother plant to microsites with physical and biological characteristics that enhance the probability of success in the germination and establishment of the plantule [19].
A number of mammals are regular frugivores and seed dispersal vectors; the ''carnivores'' (particularly bears, procyonids, some mustelids and canids) are among the most important mammalian dispersal agents. Most genera of fleshy fruits are eaten by terrestrial mammals, but few species are consumed only by mammals [20]. Some members of the order Carnivora are omnivorous, but their role as seed dispersers is still little understood [20,21]. Some species of Canidae and Procyonidae are considered efficient dispersers [20,22-27] since they consume and defecate seeds without affecting subsequent germination. In addition, these species travel relatively long distances daily and consume different types of vege s. As a result the seeds ingested can be carried far from the mother-plant, to areas where their densities are possibly lower, thereby reducing the risk of predation, attack by pathogens, intraspecific competition and inbreeding. Such dispersal provides the opportunity to colonize new areas with favorable conditions for seed germination [2,4]. In this mode of dispersal, the seeds density [28] and the number of species in a given sample [8] need to be considered as factors that can influence the success of colonization.
Despite the growing interest of fruit traits of vertebrate dispersed plants, the strategies used in plant architecture to facilitate or restrict fruit consumption and digestions by dispersers have been poorly investigated. Plant architecture can be defined as all of the parameters that permit an understanding of the plant’s structural organization, such as the internal angle of opening and the arrangement and orientation of the leaves and branches along the plant [29,30].
The B. balansae , common names Heart-of-flame (English) and gravatá and caraguatá (Portuguese), is a terrestrial bromeliad that forms a rosette about 1.5 meters high. The internal leaves form a cone of different angles, depending on the phenological state of the plant. In the period of flowering, the rosette opens exposing the inflorescence, remaining open throughout the development of the fruit. Like most vegetable families, the Bromeliaceae maintains mutualistic nonsymbiotic relationships with their dispersers, and such relationships are of great importance for maintenance of the species [4,31-33].
The objectives of this work were: 1) to identify the frugivores of B. balansae by analyzing feces and footprint traps, 2) to test the efficiency of potential dispersers of B. balansae by examining the germination of seeds that had passed through the digestive tract of the coati (N. nasua: Carnivora: Procyonidae) and the crab-eating fox (C. thous: Carnivora: Canidae) compared to control seeds collected from ripe fruits, and 3) to compare individuals of B. balansae in the vegetative and reproductive states, in order to show the difference in plant architecture.
Although the plant with ripe fruits have the open, rosette leaves still remain slightly above the ground with distances ranging from 23 to 102 cm, which could still restrict access of certain frugivorous. To check this hypothesis we compared the visitation by frugivores on fruit in the plant or placed on the ground.
Materials and Methods
Study area
This study was conducted in the Ecological Reserve of Serra do Japi (23°11′ S, 46°52′ W) in Jundiaí, state of São Paulo, southeastern Brazil. This is an area of environmental protection (APA) and consists in one of the last large areas of continuous Atlantic Rainforest in the interior of São Paulo state. It is under considerable human pressure because the extensive urbanization along its border [34,35]. The altitude of this area ranges from 700 to 1300 m and the flora is characterized by plants of the Atlantic Rainforest and also Montane Semideciduous Mesophyll Forest of the inland plateau. The mean annual precipitation is 1395 mm, with a rainy season from October to March and a dry season from April to September. The mean monthly temperature varies from 11.8°C to 15.3°C in July, and 18.4°C to 22.2°C in January, in the highest and lowest regions respectively [36]. This study was conducted during July and August of 2000. This region showed a marked seasonality, with a wet summer (December to March and a dry and cold winter (July-September) [36].
Plant species
B. balansae is a bromeliad native and not endogenous species, occurring in Colombia, Bolivia, Paraguay, Argentina and Brazil. In Brazil, it is distributed in the Amazonian, Brazilian Savanna, Atlantic Rainforest and Pantanal. This plant presenting green leaves arranged in rosette with height between 70 cm and 1.5 m with very sharp spines. In the period of flowering, the central region of the plant acquires bright red color and later white. Then, an infructescence with various orange fruits develops. Due to the placement that the rosette and flowers present, this species is known as the "heart of flame". In the local where this study was conducted, usually B. balansae fructify during the winter, corresponding to July and August. It has a sweet fruit, with white pulp and several small seeds, which is an important source of food resources for many species of animals, such as birds, mammals, reptiles and great diversity of insects [31,33,37] (Figure 1).

Figure 1: Bromelia Balansae (Bromeliaceae). Plant in reproductive stage with orange and mature fruits and open rosette with its leaves lying down, exposing the infructescence to frugivores.
Potential seed dispersers
Since most the carnivores have twilight or nocturnal habits, their observation in the wild is very difficult, making it necessary to use vestiges, such as footprints, feces and food remains, to detect their presence [38]. If correctly interpreted, these vestiges can provide a reliable identification of the species that has produced it, as well as information its ecology [39], including feeding strategies.
To register possible seed dispersers of B. balansae we installed footprints traps in 20 plants and quantified the number of ripe fruits in every one. Each trap consisted of a layer of sieved earth approximately 3 cm deep covering a circular area around the plants (Figure 2). The plots were visited in the morning and the footprints were identified in the field according to Becker and Dalponte [39]. All of the feces encountered were collected to determine the frugivores based on the marks left by the fruit consumer. The species that produced the feces were identified at the place of collection by associating the tracks with other evidence such as the consistence and appearance of the feces. When it was difficult to identify the species or when the identification was doubtful, the hair ingested during the grooming was used to complete the identification of the disperser. The hair was identified by comparing the marrow and cuticle patterns with known samples of hair, using the techniques described by Quadros [40].
Germination tests

Figure 2: Footprint trap to identify potential seed dispersers of B. balansae installed around the plant. The trap consists of a layer of sieved earth approximately 3 cm deep covering a circular area.
The footprints and feces showed coatis coati (N. nasua ), crabeating- fox (C. thous ) and Didelphis sp. (Mammalia: Marsupialia) as the mainly frugivores of B. balansae . How we have not enough seeds of B. balansae from the feces for seed germination tests, we collected ripe fruits in the field and offer them to these captive animals in a Zoo.
Seeds of B. balansae that had passed through the digestive tract of coatis (n=36 seeds) and crab-eating-foxes (n=50) were collected from the feces and used in the germination tests. Unfortunately, we did not obtain enough seeds from the feces of Didelphis sp. to be used in the germination test. For the experiment with the crab-eating-foxes we collected 50 seeds from the feces and 100 seeds from fresh ripe fruit, which were used as control. The control consisted in two levels: 1) 50 seeds of which the pulp was removed (n=50); 2) 50 seeds in which pulp was kept (n=50). Additionally, the effect of coati gut passage was determined by 36 seeds obtained from its feces, and this same seed amount was used as control. The experiments were carried until there was no germination for at least 30 consecutive days. For each treatment, all of the seeds were placed in sterile petri dishes covered with filter paper that was wetted with a fungicidal solution (0.1% Micostatin) every two days. To test for differences in germination percentage and germination time between the control group versus seeds found in the samples, we used the chi-square test [41].
Relationship between the arrangement of the leaves and the access to the fruits
To describe the architecture of vegetative and reproductive plants, we calculated the angle formed by the most internal leaves of 30 specimens of B. balansae (15 with fruits and 15 without fruits). The measurements obtained included: H1 and H2 (height of the most internal opposite leaves); and D1 and D2 (distance between the apex of these leaves and the central axis of the plant) (Figure 3). We assumed that H and D formed a right-angle triangle with the leaf so that it was possible to calculate the angles α and ß using the formula:

Figure 3: Scheme of B. balansae . H1 and H2=height of the most internal opposite leaves and D1 and D2=distance between the apex of these leaves and the central axis of the plant.
tg α=D1/H1 and tg β=D2/H2
The opening angle θ was the sum of the two angles: θ=α+β
We chose to study the internal leaves because they are the last protection barrier before reaching the fruits. The Mann-Whitney test [41] was used to compare the averages of the internal angles (θ) of plants with and without fruits, as well as the distances between opposite leaves (D1+D2). A value of p<0.05 indicated significance.
To determine if plants with open rosettes still restrict the access of certain frugivoros to ripe fruits, we installed footprint traps beneath 20 B. balansae individuals with ripe fruits. For each plant, we installed a control trap at a site approximately 30 m away. Each control trap consisted of a plot 90 cm in diameter covered by 3 cm of soil with 10 ripe fruits at the center. The footprints were recorded only when the animal reached the fruits.
Statistical analyses
To test for differences in germination percentage and germination time between the control group versus seeds found in the samples of coati and crab-eating fox, we used the chi-square test [41].
Results
Plant species
B. balansae is a terrestrial bromeliad native and the mean fruit weight was 18.89 ± 6.12 g (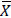 ± SD; n=10), the mean weight of peel was 12.84 ± 3.78 g ( ± SD; n = 10), the mean weight of pulp with seeds was 6.05 ± 2.51 g ( ± SD; n=10) and the mean number of seeds per fruit was 23.30 ± 9.43g ( ± SD; n=10) and the mean seed weight was 0.6 ± 0.26 g ( ± SD; n=10).
± SD; n=10), the mean weight of peel was 12.84 ± 3.78 g ( ± SD; n = 10), the mean weight of pulp with seeds was 6.05 ± 2.51 g ( ± SD; n=10) and the mean number of seeds per fruit was 23.30 ± 9.43g ( ± SD; n=10) and the mean seed weight was 0.6 ± 0.26 g ( ± SD; n=10).
Identification of possible dispersers
We recorded a total of 52 footprints and identified crab-eating foxes (n=13), coatis (n=4) and opossums, Didelphis sp. (n=12) as potential dispersers. We considered these three species to be possible dispersers because they took whole fruits from the plant at each visit. We also identified birds (n=11) and rodent (n=4) tracks, but since the fruits were only partially eaten we did not consider them as dispersers. Some poorly defined tracks were not identified (n=8) (Table 1).
Of 22 feces collected only five contained B. balansae seeds: three were from crab-eating-foxes and two from coatis. The other food items in the feces were not identified or quantified and feces that did not contain B. balansae vestiges were discarded. In the three feces of crabeating- foxes, we found 6, 9 and 24 seeds each, while those of the two coatis contained 9 and 11 seeds each. No seeds from other species were found in these feces (Table 1).
| Species | Common name | FP | FE (range) | Feces | Seeds (range) |
|---|---|---|---|---|---|
| Didelphis sp. | Opossum | 12 | 1.6 (1-3) | 0 | - |
| C. thous | Crab-eating fox | 13 | 2.2 (1-4) | 3 | 13 (6-24) |
| N. nasua | Coati | 4 | 2.0 (1-4) | 2 | 10 (9-11) |
| Undetermined | Rodents | 4 | * | 0 | - |
| Undetermined | Birds | 11 | * | 0 | - |
| Undetermined | Undetermined | 8 | - | - | - |
| Total | 52 | - | 5 | - |
FP:Total number of registered footprints; FE:Average of fruits eaten by visit; *: fruits that were partially eaten; Fec: Number of feces found that contained B. balansaeseeds; Seeds: Number of B. balansae seeds found in the feces.
Table 1: Potential seed dispersers of B. balansae .
Germination test
Coatis
The experiment with N. nasua lasted for 96 days and the seeds required 41 days to germinate and once again, seeds without pulp germinated more frequently than seeds with pulp: all of the seeds recovered from feces germinated compared to 66% of those collected from fresh fruits and with the pulp removed, and 13.8% of germination of those with pulp.
Seeds that passed through the digestive tract of N. nasua germinated faster than seeds from fruits (Figure 4). The germination of seed from coati feces started the germination five days after the beginning of experiment, while seeds from fruits germinated with 15 days of experiment.

Figure 4: Seed germination under treatment of coati gut passage and seeds that did not pass through the digestive tract.
There was germinability of 100% registered for intact seeds found in the feces of N. nasua . Thus, seeds that remained intact after gut passage showed was significantly higher germinability than that both groups of seeds, without pulp (χ2=20.4819; df=1; p=0.000006) and with pulp (χ2=75.4386; df=1; p=0.00000). In addition, the germinability of seeds with pulp removed was significantly higher than that of group of seeds with pulp kept (χ2=4.19309; df=1; p=0.040589) (Figure 5).

Figure 5: Experiment comparing the effect two treatments on the seed germination: 1) coati gut passage (seeds collected from coati feces); 2) from mature fruits (seeds with pulp removed); 3) from mature fruits (pulp kept). Significant differences in the total percentages of the germinated seeds are indicated by different letters (Chi-square test; α=0.05).
Crab-eating foxes
The B. balansae seeds germinated after 25 days and germination continued for 30 days. The experiment was finished after 84 days.
Seeds that passed through the digestive tract of C. thous germinated faster than seeds from fruits (Figure 6). In turn, seeds that remained intact after gut passage was significantly higher than that of seeds with pulp (χ2=6.63650; df=1; p=0.009), but showed no significant differences in their germinability when compared with the seeds with removed pulp (χ2=2.17, df=1, p=0.14). In addition, seed with pulp removed showed no significant differences in their germinability when compared with the group of seeds with pulp kept (χ2=0.287807; df=1; p=0.59) (Figure 7).

Figure 6: Seed germination under treatment of crab-eating foxes gut passage and seeds that did not pass through the digestive tract.

Figure 7: Experiment comparing the effect two treatments on the seed germination: 1) fox gut passage (seeds collected from fox feces); 2) from mature fruits (seeds with pulp removed); 3) from mature fruits (pulp kept). Significant differences in the total percentages of the germinated seeds are indicated by different letters (Chi-square test; α=0.05).
Crab-eating fox vs. Coati vs. fruit
In short, our data indicated that the coati gut passage has higher positive effect on the seed germination of B. balansae seeds than fox and fruit treatments. On the other hand, fox treatment presented higher seed germination than seeds from mature fruits without pulp (Figure 8). In turn, germinability of seeds from coati feces was significantly higher than that from fox feces (χ2=38.0952; df=1; p=0.000000) and also than seeds from fruits (χ2=68.4564; df=1; p=0.000000). Finally, seeds that remained intact after fox gut passage showed significantly higher germinability compared with the seeds from fruits (χ2=7.43487; df=1; p=0.006397).

Figure 8: Cumulative percentages of germinated seeds over a 56- days period for the B. balansae seeds extracted from mature fruits and from C. thous (coati) and N. nasua (fox) feces. Significant differences in the total percentages of the germinated seeds are indicated by different letters (Chi-square test; α=0.05).
Relationship between the arrangement of the leaves and the access to the fruits
The average value of the opening angle (θ) of the B. balansae , and the average distance among the more internal leaves (D1+D2), were significantly greater in plants with ripe fruits (Table 2).
| Parameter | Without fruits (n=15) | With fruits (n=15) | |||
|---|---|---|---|---|---|
| ± SD |
range | ± SD |
range | ||
| H1 (cm) | 109.1+19,3 | 76-147 | 45.7+20.1 | 23.0-102.0 | |
| H2 (cm) | 125.1+18,2 | 97-156 | 84.4+38.9 | 38-186 | |
| D1 (cm) | 23.5+17,5 | 8-77 | 56.1+20.2 | 11-92 | |
| D2 (cm) | 34.0+18,7 | 8-82 | 87.4+33.6 | 21-140 | |
| D1+D2 (cm) | 57.5+37,2 | 16-159 | 143.5+53.8 | 32-232 | |
| α (°) | 12.1+8,0 | 4.2-35.6 | 78.5+38.6 | 18.4-179.4 | |
| b (°) | 15.0+7,8 | 3.7-28.2 | 46.8+15.8 | 12.7-64.6 | |
| Θ (α+β) | 27.1+13,8 | 7.9-54.7 | 125.2+44.4 | 66.6-227.1 | |
Table 2: Relationship between the arrangement of the leaves and exhibition of the fruits.
There was no significant difference in the number of footprints of potential large dispersers in the isolated (control) traps compared to traps around the plants (Table 3) (Mann-Whitney test, p=0.699). In contrast, the footprints of small rodents and birds were seen only in control traps.
| Recorded specie | Control (traps without plant) | Plants (plot around) |
|---|---|---|
| C. thous | 16 | 15 |
| N. nasua | 18 | 21 |
| Didelphis sp. | 9 | 7 |
| Unidentified rodents | 7 | 0 |
| Unidentified birds | 11 | 0 |
| Total | 61 | 43 |
-The tracks were recorded only when the fruits were reached.
Table 3: Occurrence of footprints around and distant from B. balansae plants.
Discussion
Most of the Procionidae and South American Canidae are opportunists and generalists, with fruits, invertebrates and small vertebrates being the principal components of their diet [22,42-51]. In our study the mainly frugivores of B. balansae were coatis and crabeating foxes, and in agreement with Rocha [52], they were considered true dispersers since the seeds passed intact through the digestive tract and were deposited far away from the mother-plant. On the other hand, birds and some non-identified rodents were considered potential seed dispersers since they fed on the pulp of the fruit, leaving the seeds close to the mother-plant. Another non-identified rodent was considered seed predators since they damaged the seeds when they fed on the fruits.
Our data showed that the seed germination performance (final percentages and speed) increased significantly after passing through the N. nasua and C. thous guts compared with those of the seeds from mature fruits. Thus, seed passage through the digestive tracts of coatis and foxes shortened seed dormancy corroborating innumerous other studies to other vertebrate frugivorous (e.g., Varela et al. [53]; Vasconcellos-Neto et al. [29]; Alves-Costa et al. [54]). Varela et al. [53] discussed that in treatments that are free of pulp, the faster germination may be consequences of the seed coat characteristics and of the digestive process of vertebrates that may reduce the thickness of the seed coats or stony endocarp, thus increasing permeability to water and gases.
Here authors showed that the removal of pulp also increases the germination rates of B. balansae seeds in comparison of those seeds with pulp. According to Barnea et al. [55], Traveset et al. [13], Varela et al. [53] and Ribeiro et al. [56], removal of the pulp increases germination by reducing the growth of fungi and due the removal of chemical inhibitors present in the fruit pulp, or mechanical or chemical scarification of the seed coat, which increases permeability to water and gases. In our study, none of the seeds recovered from the feces had any pulp and showed faster germination and higher germinability, corroborating this theory above discussed.
In this context, our results for N. nasua confirm that the seed passage through the digestive tracts affects positively the seed germination performance (final percentages and speed) increased significantly after passing through the coatis’ guts compared with those of the seeds from fruits, consistent with data from other studies [57,58]. On the other hand, Alvez-Costa et al. [54] studying consumption and seed germination of 53 fruit species, also in an Atlantic Rainforest in southeastern Brazil, verified that the ingestion of seeds by coatis did not alter speed or success of seed germination for any of the species studied, except for one.
Thus, although our data indicate a positive effect of gut passage of B. balansae seeds through the coati digestive tract, recent studies have found the opposite results, evidencing that the comprehension of all features that affect the germination performance depends on the type and intensity of ecological interaction between the frugivore and plant species and involves a complexity of features as seed morphology and physiology, chemical substances in the pulp and countless others.
In contrast, Ribeiro et al. [56] studying Melastomataceae spp., suggested that gut passage has null effect on the seed germination and that the crucial the removal of the seed pulp. Second to him, the seed cleaning is important to remove germination inhibitors in the fruit pulp of the species studied. The experiment with C. thous agrees with Ribeiro et al. [56], in which we verified greater germinability in the seeds obtained from mature fruits and whose pulp was manually removed than for gut-passed seeds, and the lower germination rate was observed for seeds with pulp obtained from ripe fruits. In fact, our data strongly indicate that the C. thous gut passage has a negative effect when compared to the germinability of seeds obtained from mature fruits with manually removed pulps.
In sum, our data for N. nasua indicate positive effect of gut passage of B. balansae seeds, whereas data for C. thous suggest that the most important for the germination performance is the removal of the pulp and that the passage through the digestive tract had a negative effect on germination, evidenced by the fact that seeds with pulp manually removed had a higher germination rate than those from fox feces. These contrasting data showing that the understanding the complexity of factors that affect the outcome of frugivores gut passage on seed germination is a challenging theme in seed dispersal ecology. Actually, as discussed by Traveset et al. [59] even after decades of studies, there is still no consensus regarding which specific mechanism is the main responsible for changes in germination rate and time following gut passage.
The Atlantic forest is highly fragmented. Especially in the state of São Paulo, about 70% of protected forest fragments are <1000 ha and 80% of unprotected fragments are <50 ha [60]. The Thus, the size of the Ecological Reserve of Serra do Japi is similar to several other Atlantic Rainforest remnants. In fragments like these, the residual large-bodied frugivores are primarialy the Rusty-margined Guan (Penelope superciliaris ) [61] and the mammals Didelphis spp., Callithrix spp., C. thous and N. nasu [62]. Therefore, in fragments such as those found in Ecological Reserve of Serra do Japi large-seeded and/or large-fruited plant species may have few frugivores species as seed dispersers. As suggested by Alves-Costa et al. [54], the seed dispersal services provided by N. nasua are more similar to those provided by C. thous , because both are dispersers of plant species with similar seed size and fruit colour, and also, both frugivores have the potential to disperse seeds over large distances.
The fruiting period of most plant species in Ecological Reserve of Serra do Japi is comprehended between at the end of the dry season and beginning of the wet season (October to December) [9]. Thus, the availability of B. balansae fruits in the beginning of the dry season (June to August) period of resource scarcity consists in an important ecological strategy that allows generalists species as coatis and crabeating foxes use its fruits as food source and consequently, act as important seed dispersers of this bromeliad species.
Vegetative individuals of B. balansae shows small internal leaves angles, and how the leaves are hard and with many thorns in its margins they could protected the central gem were the plant will produce the inflorescence. Additionally, flowering and fruiting individuals of B. balansae show open rosettes to facilitate the access of floral visitors and frugivorous. However, even when the fruits were exposed, the morphology of the leaves (hard, with many thorns) and plant architecture hindered the access of small animals, such as small rodents and birds. Thus, the natural characteristics of the plant favored the access of larger dispersers as coatis and crab-eating foxes which, as suggested by Alves-Costa et al. [54] have potential to promote seed dispersion over long distances.
Acknowledgements
We thank the staff at the Base for Ecological Studies and Environmental Education "Miguel Gastarde", and the City Hall of Jundiaí for providing the necessary infrastructure and Dr. Woodruff Whitman Benson for guidance during this work. We also thank the “Quinzinho de Barros” Municipal Zoological Park in Sorocaba, especially Cecília Pessutti and Eugênio José da Silva, for their help with the germination tests. Paulino-Neto HF was supported by a grant from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, grant no. 724/00), E. Nakano-Oliveira (CAPES, grant no. 724/2003), M.M.A. Jardim (FAPESP no. 99/00024-8), J. Vasconcellos- Neto (CNPq, grant no. 300539/94-0). Paulino-Neto thanks Milene Souza Rodrigues Paulino (my wife) for unconditional support during all phases of this study and Otávio Rodrigues Paulino (my son) for the contagious joy and inspiration to always move on.
References
- Medellin RA, Gaona O (1999) Seed dispersal by bats and birds in forest and disturbed habitats of Chiapas, México. Biotropica 31: 478-485.
- Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annu Rev EcolSyst 13: 201-228.
- Gautier-Hion A, Duplantier JM, Quris R, Feer F, Sourd C (1985) Fruit characteristics as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community. Oecologia 65: 324-337.
- Howe HF, Westley LC (1988) Ecological relationships of plants and animals. Oxford University Press. Oxford. p: 273.
- Fleming TH, Breitwisch R, Whitesides GH (1987) Patterns of tropical vertebrates frugivore diversity. Annu Rev EcolSyst 18: 91-109.
- Fleming TH, SOSA VJ (1994) Effects of nectarivorous and frugivorous mammals on reproductive success of plants.J Mammal 75: 845-851.
- Laman TJ (1996) Ficus seed shadows in a Bornean rain forest. Oecologia 107: 347-355.
- Loiselle BA (1990) Seeds in droppings of tropical fruit-eating birds: importance of considering seed composition. Oecologia 82: 494-500.
- Morellato LPC, Leitão-Filho HF (1992) Patterns of fruiting and dispersal in the Serra do Japi. In: Morellato LPC (org.). Natural History of the Serra do Japi. Ecology and Preservation of a Forest Area in Southeast Brazil. Ed. Unicamp, Campinas. p: 321.
- Izhaki I, Safriel UN (1990) The effect of some Mediterranean scrubland frugivores upon germination patterns. J Ecol 78: 56-65.
- Bodmer RE (1991) Strategies of seed dispersal and seed predation in Amazonian ungulates. Biotropica, 23: 255-261.
- Agami M, Waisel Y (1986) The role of mallard (Anasplatyrhynchos) in distribution and germination of Najas marina L. seeds. Oecologia 68: 473-475.
- Traveset A, Willson MF (1997) Effects of birds and bears on seed germination of fleshy-fruits plants in a temperate rainforest of southeast Alaska. Oikos 80: 89-95.
- Samuel IA, Levey DJ (2005) Effects of gut passage on seed germination: do experiments answer the questions they ask? FunctEcol 19: 365-368.
- Rodriguez-Perez J, Riera N, Traveset A (2005) Effect of seed passage through birds and lizards on emergence rate of mediterranean species: Differences between natural and controlled conditions. FunctEcol 19: 699-706.
- Janzen DH (1981) Digestive seed predation by a Costa Rican Baird’s tapir. Biotropica 13: 59-63.
- Izahaki I, Korine C, Arad Z (1995) The effect of bat (Rousettusaegyptiacus) dispersal on seed germination in eastern Mediterranean habitats. Oecologia 101: 335-342.
- Rowell TE and Mitchell BJ (1991) Comparison of seed dispersal by guenons in Kenya and capuchins in Panama. J Trop Ecol7: 269-274.
- Stiles EW, White DW (1986) Seed deposition patterns: influences of season, nutrients, and vegetation structure. In: Estrada A, Fleming TH (eds) Frugivores and seed Dispersal. Dr. W. Junk Publishers, Dordrecht. pp: 45-54.
- Willson MF (1993) Mammals as seed-dispersal mutualists in North America. Oikos 67: 159-176.
- Herrera CM (1989) Frugivory and seed dispersal by carnivorous mammals, and associated fruit characteristics, in undisturbed Mediterranean habitats. Oikos. 55: 250-262.
- Motta-Junior JC, Lombardi JA, Talomoni AS (1994) Notes on crab-eating fox (Dusicyon thou) seed dispersal and food habits in southeastern Brazil.Mammalia 58: 156-159.
- Gompper ME (1996) Sociality and asociality in white-nosed coatis (Nasuanarica): foraging costs and benefits. Behavioral Ecology 7: 254-263.
- Gompper ME (1997) Population ecology of the white nosed coati (Nasuanarica) on Barro Colorado Island, Panama. J ZoolLond 241: 441-455.
- Wilson JA, Barry T (1999) Diet and seed dispersal efficiency of the gray fox (Urocyoncinereoargenteus) in Chaparral. South. Calif. Ac. of Science 98: 119-126.
- Cheida CC (2002) Diet, seed dispersal and foraging behavior of Cedarhorn (Carnivora: Canidae) in an Atlantic Forest area: Salto Morato Natural Reserve, Guaraqueçaba, Paraná. Monograph, Center of Biological Sciences, State University of Londrina. p: 70.
- Rocha VJ, Reis NR, Sekiama NL (2004) Diet and seed dispersal by Cerdocyonthous (Linnaeus) (Carnivora: Canidae) in a forest fragment of Paraná, Brazil. Rev Bras Zool 21: 871-876.
- Augspurger CK, Kitajima K (1992) Experimental studies of seedling recruitment from contrasting seed distributions. Ecol73: 1270-1284.
- Vasconcellos-Neto J, De Albuquerque LB, Silva WR (2009) Seed dispersal of SolanumthomasiifoliumSendtner (Solanaceae) in the Linhares Forest, Espirito Santo state, Brazil. Acta Bot Bras 23: 1171-1179.
- Paulino-Neto HF, Nakano-Oliveira E, De AssisJardim MM, Medeiros RP,Vasconcellos-Neto J, et al. (2013) Frugivory by Sturniralilium bats (Phyllostomidae) on Solanummauritianum (Solanaceae) in southeastern Brazil. Studies on Neotropical Fauna and Environment 48: 183-189.
- Durigan G, Baitello JB, Franco GADC and Siqueira MF (2004) Plants of CerradoPaulista: images of a threatened landscape. Pages & Lyrics Editora e Gráfica. Sao Paulo. p: 475.
- Fontoura T, Cazetta E, Nascimento W, Catenacci L, De Vleeschouwer K, et al. (2010) Diurnal frugivores on the BromeliaceaeAechmeadepressa L.B. Sm. from Northeastern Brazil: the prominent role taken by a small forest primate. Biota Neotrop 10: 351-354.
- Flora of Brazil (2020) Under Construction. Botanical Garden of Rio de Janeiro.
- Morellato LPC (1992) Seasonality and dynamics of forest ecosystems in the Serra do Japi. In: Morellato LPC (Org) Natural History of the Serra do Japi. Ecology and Preservation of a Forest Area in Southeast Brazil. Ed. Unicamp, Campinas. p: 321.
- Polli PR, Vasconcellos-Neto J (2012) Environmental Impacts in the Serra do Japi. In: Vasconcellos-Neto J, Polli PR, Penteado-Dias AM (Orgs.) New Looks, New Knowledge About the Serra do Japi: Echoes of its biodiversity. Publisher CRV. Curitiba. pp: 545-583.
- Pinto HS (1992) Climate of the Serra do Japi. In: Morellato LPC (ed) Natural History of the Serra do Japi. Ecology and Preservation of a Forest Area in Southeast Brazil. Unicamp, Campinas. pp: 30-39.
- Pott A, Pott VJ, Sobrinho AAB (2004) Plants useful for survival in the Pantanal. In: Symposium on Natural and Socio-Economic Resources of the Pantanal, 4. Corumbá.
- Pardini R, Ditt EH, Cullen L, Bassi C, Rudran R (2003) Rapid survey of medium and large terrestrial mammals. In: Cullen JrL, Rudran R, Valladares-Padua C (Orgs)Methods of studies in conservation biology and wildlife management. Curitiba: Publisher of the Federal University of Paraná and Boticário Foundation for Nature Protection. pp: 181-201.
- Becker M, Dalponte JC (1991) Stars of Brazilian Wild Mammals-a Field Guide. Edund
- Quadros J (2002) The use of hair in the identification of Brazilian mammals. Doctoral thesis, Federal University of Paraná (UFPR) Curitiba.
- Zar JH (1996) Biostatistical Analysis. Prentice Hall. New Jersey.
- Berta A (1987) Origin, diversification, and zoogeography of the South American Canidae.In: Patterson, B.D. and Timm, R.M. (eds.) Studies in NeotropicalMammalogy: Essays in Honor of Philip Hehkowitz. FieldianaZool 39: 455-471.
- Emmons LH, François F (1990) Neotropical Rainforest Mammals: a Field Guide. University of Chicago Press.
- Nowak RM, Paradiso JL (1991) Walker’s Mammals of the World. 5th ed. Baltimore. The Johns Hopkins University Press 2.
- Facure KG, Giaretta AA (1996) Food habits of carnivores in a coastal Atlantic forest of southeastern Brazil. Mammalia 60: 499-502.
- Alvez-Costa CP (1998) Frugivory and seed dispersal by quatis (Nasuanasua) in the Mangabeiras Park, Belo Horizonte, MG. Master's dissertation, Institute of Biology, State University of Campinas (UNICAMP).
- Nakano-Oliveira E (2002) Food ecology and living area of carnivores of the Ipanema National Forest, Iperó, SP (Carnivora: Mammalia). Master's dissertation Institute of Biology, State University of Campinas (UNICAMP).
- Bueno AD, Motta-Junior JC (2004) Food habits of two syntopiccanids, the maned wolf (Chrysocyonbrachyurus) and the crab-eating fox (Cerdocyonthous), in southeastern Brazil. Rev ChilHist Nat 77: 5-14.
- Cheida CC, Nakano-Oliveira E, Fusco-Costa R, Rocha-Mendes F, Quadros J et al. (2006) OrdemCarnivora. In: Reis NR, Peracchi, AL, Pedro WA, Lima IP (eds) Mammals of Brazil. Londrina: REIS N R p: 437.
- Nakano-Oliveira E (2006) Ecology and Conservation of Carnivorous Mammals of the Atlantic Forest in the Lagunar Estuary Complex of Cananéia, State of São Paulo. Doctoral thesis Institute of Biology, State University of Campinas, UNICAMP.
- Gatti A, Bianchi R, Rosa CRX, Mendes SL (2006) Diet of two sympatric carnivores, Cerdocyonthous and Procyoncancrivorus, in a restinga area of Espirito Santo State, Brazil. J Trop Ecol 22: 227-230.
- Rocha VJ (2001) Ecology of mammals of medium and large size of Mata dos Godoy State Park, Londrina, PR Doctoral thesis, Federal University of Paraná (UFPR) Curitiba. p: 124.
- Varela O, Bucher EH (2006) Passage time, viability, and germination of seeds ingested by foxes. J Arid Environ 67: 566-578.
- Alvez-Costa CP, Eterovick PC (2017) Seed dispersal services by coatis (Nasuanasua, Procyonidae) and their redundancy with other frugivores in southeastern Brazil. ActaOecologica 32: 77-92.
- Barnea A, Yo-Toy Y, Friedman J (1991) Does ingestion by birds affect seed germination? FunctEcol 5: 394-402.
- Ribeiro RC, Figueiredo MLN, Picorelli A, Oliveira DMT, Silveira FAO (2016) Does seed coat structure modulate gut-passage effects on seed germination? Examples from Miconieae DC. (Melastomataceae). Seed Sci Res 26: 139-147.
- Carvalho-Ricardo MC, Uieda W, Fonseca RCB, Rossi MN (2014) Frugivory and the effects of ingestion by bats on the seed germination of three pioneering plants. ActaOecol 55: 51-57.
- Giordani L, Baraza E, Camargo-Ricalde SL, Moe SR (2014) The domestic goat as a potential seed disperser of Mimosa luisana (Leguminosae, Mimosoideae) in the Tehuacán-Cuicatlán Valley, Mexico. J Trop Ecol 31: 91-94.
- Traveset A, Rodríguez R, Pías B (2008) Seed traits changes in dispersers’ gut and consequences for germination and seedling growth. Ecology 89: 95-106.
- Ditt EH (2002) Forest fragments in Pontal do Paranapanema. Environment 207.
- Pizo MA (2004) Frugivory and habitat use by fruit-eating birds in a fragmented landscape of southeast Brazil. OrnitolNeotrop 15: 117-126.
- Gheler-Costa C (2002) Non-flying mammals of the ''Luís de Queiróz'' Campus, University of São Paulo, Piracicaba, State of São Paulo. Master dissertation, ESALQ. University of São Paulo, Piracicaba, SP, Brazil.
Relevant Topics
- Aquatic Ecosystems
- Biodiversity
- Conservation Biology
- Coral Reef Ecology
- Distribution Aggregation
- Ecology and Migration of Animal
- Ecosystem Service
- Ecosystem-Level Measuring
- Endangered Species
- Environmental Tourism
- Forest Biome
- Lake Circulation
- Leaf Morphology
- Marine Conservation
- Marine Ecosystems
- Phytoplankton Abundance
- Population Dyanamics
- Semiarid Ecosystem Soil Properties
- Spatial Distribution
- Species Composition
- Species Rarity
- Sustainability Dynamics
- Sustainable Forest Management
- Tropical Aquaculture
- Tropical Ecosystems
Recommended Journals
Article Tools
Article Usage
- Total views: 3702
- [From(publication date):
December-2016 - Aug 29, 2025] - Breakdown by view type
- HTML page views : 2805
- PDF downloads : 897