Research Article Open Access
Effects of Climate Change on Invasion Potential Distribution of Lantana camara
| Neena Priyanka* and Joshi PK | |
| Department of Natural Resources, TERI University, New Delhi, India | |
| Corresponding Author : | Neena Priyanka Department of Natural Resources TERI University, New Delhi, India E-mail: neenapriyanka@gmail.com |
| Received August 05, 2013; Accepted September 20, 2013; Published September 28, 2013 | |
| Citation: Priyanka N, Joshi PK (2013) Effects of Climate Change on Invasion Potential Distribution of Lantana camara. J Earth Sci Clim Change 4:164. doi:10.4172/2157-7617.1000164 | |
| Copyright: © 2013 Priyanka N, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. | |
Visit for more related articles at Journal of Earth Science & Climatic Change
Abstract
Climate change appears to be affecting global patterns of invasive species distribution. Forecasts based on ecological niche modeling suggest that greater impacts can be expected in the future. However, such projections are contingent on assumptions regarding the future climate conditions and invasion potential of a species. This study explores the relationship between climate change and potential distribution of Lantana camara in the National Parks of Jim Corbett and Rajaji (Uttarakhand, India). Using three representative climate change models viz., CSIRO (Commonwealth Scientific and Industrial Research Organization), CCCMA (Canadian Centre for Climate Modeling and Analysis) and HadCM3 (Hadley Centre for Climate Prediction and Research’s General Circulation Model) across the time slices 2020 to 2080 under two regional climate change scenarios A2a and B2a, Lantana camara potential distribution models were derived. The model projections were in consensus that invasion range was likely to expand and infestation would be more severe under the A2a scenario indicating that the species may prefer warmer conditions. Taken together, the modeled results suggest that in the future, the two National Parks may be impacted largely by the gregarious presence of Lantana camara. Predictive models can provide resource managers with a tool for the early detection of invasive species and help circumvent negative ecological impacts resulting in substantial economic savings.
| Keywords |
| Lantana camara; Niche modeling; Climate change; Invasion; National parks |
| Introduction |
| Invasive plants have altered the biodiversity and ecological integrity of native habitats [1,2]. They have affected natural processes [3], homogenized flora [4], caused the extinction of species [5,6], compromised agriculture production [7], and damaged ecosystem resources [8,9]. The problem arises because questions of when and why some invasive species escape and cause nuisance remain unanswered [10]. The problem has been further exacerbated by a considerable climate change in the natural ecosystems. Climate change may be described as a long-term shift in the regional trends of weather measured by changes in its rate, range and magnitude [11]. It has been plaguing much of the world and is a serious concern at continental, regional, national and local scales [11]. |
| The relationship between species invasion and climate change is complex and linked in several ways. Climate change is likely to enhance the dimensions of invasive to occupy new areas, while simultaneously increasing the adaptability in natural communities by disturbing the dynamic equilibrium maintaining them [12-14]. On the other hand, invasive species influence the magnitude, rate and impact of climate change by altering ecosystem composition, structure and function [15-17]. The Intergovernmental Panel on Climate Change (IPCC) covers the issue in considerable detail and reports that the recent worldwide warming will result in pole ward and altitudinal shifts in plant ranges. The report also forecasts that many ecosystems will become vulnerable to biological invasions as climatic barriers are removed. The IPCC concludes that an increase of greater than 1.5°C-2.5°C in the global average temperature will cause dramatic changes in species distribution and ecosystem function, resulting in overwhelmingly negative consequences for ecosystem sustainability [11]. |
| Thus, in order to improve our understanding of likely impacts of invasion of a given species in the wake of climate change, knowledge of the current presence or absence and likelihood of its establishment or spread under future climate change scenarios is of the utmost importance. |
| Climate scenarios are the alternative images of how the climate in the future may unfold. The Special Report Emissions Scenarios (SRES) describes four climate scenario “families”, each with a divergent future description. Since it is difficult to project far-off future emissions and other human factors that influence climate, scientists use a range of scenarios making various assumptions about economic, social, demographic, technological, and environmental conditions to project future global warming (Table 1). Scenarios range from Low emissions (B1, B2: less pronounced future warming than A1 and A2) to High emissions (A1 and A2) in three time slices: ‘2020s’ (2010-2039), ‘2050s’ (2040-2069) and ‘2080s’ (2070-2099). |
| To predict the potential distribution of invasive species under the different climate change scenarios, various modeling algorithms have been used by researchers [18-21]. Here, the choice of the modeling algorithm plays a critical role in determining the accuracy of the predictions. Most algorithms require both presence and absence data sets. This is difficult to collate because while the presence data can be collected with confidence, the absence data has high inherent uncertainties [18,22]. Hence, modeling methods that require presence only dataset, and can predict effectively even with limited available data have been used more commonly [21,23]. |
| The aim of the present study is to assess through modeling the future distribution of the robust and gregarious invasive species-Lantana camara - relative to its current distribution potential in the Western Himalaya regions of India under different climate change scenarios. The Maxent modeling technique has been employed for assessing the current distribution. In addition, three other models-CSIRO (Commonwealth Scientific and Industrial Research Organization), CCCMA (Canadian Centre for Climate Modeling and Analysis) and HadCM3 (Hadley Centre for Climate Prediction and Research’s General Circulation Model)-have been employed to predict the future distribution range of Lantana. |
| Materials and Methods |
| Study Area |
| The study area comprises a part of the Western Himalayas including two important National Park (NP) viz. the Jim Corbett and Rajaji. The Corbett NP (29°50'-30°20' N and 77°50'-78°30' E) covering an area of 1318.54 sq. km, lies in two districts- Nainital and Pauri Garhwal of Uttarakhand, India. The park is situated at an altitudinal range of 385-1100 m absl with a temperature range of 4°C-42°C and rainfall of 1400-2800 mm. The Rajaji NP (29°56'- 30°20' N and 77°55'-79°80' E) covers an area of 820.42 sq. km and is located between Haridwar and Dehradun districts of Uttarakhand. The park is situated at an altitudinal range of 300-1,345 m absl with a minimum temperature around 13.1°C and maximum temperature around 38.9°C and average rainfall of 1200 mm. The spatial coverage of this study was deliberately made larger (a section of the Western Himalayan region was considered instead of the geographical areas of two parks) than the actual range occupied by Lantana camara to ensure encompassing the full extent of the predicted range (Figure 1). Both the parks are engrossed in the Shivalik ecosystem and the beginning of the vast Indo-Gangetic plains, thus representing vegetation of several distinct zones and forest types and diverse flora and fauna. |
| This region encircling two important national parks was selected for study because of its gregarious Lantana camara presence, high ecological and economic value, and its charisma of conservation significance, which houses large populations of huge mammals like the Indian Tiger, Asian Elephant and Swamp Deer among other important medicinal and endemic variety of plant and animal species. High human density in and around forests has resulted in strong gradients of degradation and anthropogenic pressures from the forest edge to the interior, thus paving the way for invasion by Lantana camara. |
| Input datasets |
| A total of 137 occurrence records of Lantana camara were obtained from native locations of Jim Corbett and Rajaji NP through field survey. The establishments of these occurrence records were validated from park officials and by electronic database searches such as Google, Scopus, Science Direct and published works. Current climate and scenarios of future projections (IPCC 3rd assessment data) datasets at a spatial resolution of 1 km were downloaded from the WorldClim database [24]. Three climate-change models CCCMA (Canadian Centre for Climate Modeling and Analysis), CSIRO (Commonwealth Scientific and Industrial Research Organization) and HadCM3 (Hadley Centre for Climate Prediction and Research’s General Circulation Model), for the period of 2020-2080 including two scenarios A2a and B2a were used for building invasion potential distribution model of Lantana camara. Bioclimatic layers were not available for future datasets, so these were computed from temperature and precipitation datasets and equations available at Worldclim resources (http://www.worldclim.org/). Climatic envelopes have information on temperature, precipitation, its seasonality and deviations and thus impact species distribution greatly. Besides exerting direct physiological effects on species, climatic variables also reflect energy and water availability which in turn impinges ecosystem productivity. Topographical variables such as altitude, slope, aspect, flow direction, flow accumulation and compound topographic index (CTI), have an indirect effect in influencing microclimates within species ranges [25], so these were included in present study. CTI was computed using equation 1: |
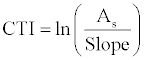 (1) (1) |
| Where, As = Flow accumulation obtained from elevation after usual pre-processing to fill sinks and find flow directions. Slope=Slope percent derived from altitude. |
| Monthly potential evapo-transpiration (PET) was calculated using Thornthwaite equation [26,27] as illustrated in equation 2. |
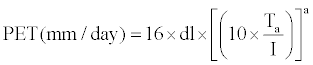 (2) (2) |
| where, |
| Ta = Mean monthly temperature (°C) |
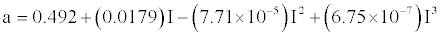 |
| dl=Day length in hours/12z |
I =S i (Annual heat index) and i=Monthly heat index given by 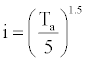 |
| These predictor variables were selected based on biological relevance to species distribution and information obtained from other spatial modeling studies carried out on Lantana camara [6,28-30]. To enable projections of future species invasion potential ranges, environmental predictors were required to be in pairs, one for current and other for future conditions. Thus, only those variables that existed for future climate scenarios and matched with variables of current climate scenarios were selected. Since topographical variables are not subject to change in future these were assumed to be constant for both time periods due to high uncertainties in projecting their future conditions. The predictor variables were subjected to multi-collinearity diagnostics using Pearson product-moment correlation coefficient to reduce data dimensionality. The correlation coefficient values range between +1 and -1 inclusive where +1 in the case of a perfect positive (increasing) linear relationship, -1 in the case of a perfect decreasing (negative) linear relationship, and some value between -1 and 1 in all other cases, indicating the degree of linear dependence between variables. In present study, Pearson value threshold of ± 0.80 was considered as elimination criteria for selecting unique variables. Amongst 73 variables obtained, numbers of predictor variables were reduced to 23 based on Pearson correlation test. |
| Modeling |
| The analysis was conducted using Maxent (http://www.cs.princeton.edu/~schapire/maxent/). The Maxent (Maximum Entropy) modeling algorithm developed by Phillips [21] is reliable even when data availability or reliability is less. It can handle both continuous and categorical variables, incorporating interactions, and modeling non-linearities [23,31,32]. It has been used in developing ecological models for various biodiversity applications [20,33,34]. Several studies have demonstrated the reliability of Maxent [4,32,35,36]. Maxent is extensively being used by the Australian government agencies to predict the distribution of key weed species under future climatic conditions [10,13,37-39]. Thus, in the current research Maxent modeling technique was used. |
| Default values for the convergence threshold (10^-5) and maximum number of iterations (1000) were selected for the model settings as suggested by Phillips [21]. Regularization multiplier was set to 1 and the “fade by clamping” option was used to control extrapolation beyond the range of the training dataset. The linear, quadratic, product, hinge and threshold functions were used. Setting of regularization values, which address the problems of over-fitting, and the selection of ‘features’ were performed automatically by Maxent as per the default rules depending upon the number of distribution records and features used in model construction. One third of the distribution records were retained at random for model evaluation using Area under Curve (AUC) of the Receiver Operating Characteristic (ROC) plot of sensitivity vs. 1-specificity [21]. Models found to have good predictive performance have test AUC value greater than 80%. Models were projected from the present to the future climate scenarios. The Jackknife test for variable importance was carried out to determine predictive ability of the variables used and their contribution to species expansion or contraction [21]. |
| Results |
| Spatial invasion potential distribution of Lantana camara |
| Using 137 occurrence localities of Lantana camara and 23 predictor variables, models were generated for depicting the potential distribution of Lantana camara under the current and the future climate change projections. |
| Current climate projections: The Maxent model depicted high infestation of Lantana camara in the Western Himalayan foothills enclosing Haridwar, Pauri Garhwal, Dehradun, parts of Tehri Garhwal and Almora regions. The species was found to be predominant in regions with altitude less than 2000 m absl, mean annual precipitation ranging from 800 to 2000 mm, annual temperature regime of 10 to 30°C, and soil ranging from sandy to gravelly and clayey loams. Lantana camara populations were found to be more in the northern and western limits as compared to the southern and eastern regions. The region in and around Jim Corbett and Rajaji NP were highly infested with the species which has already reached nuisance status. Most regions of the Jim Corbett NP manifested very high potential distribution with infestations severe in the Jhirna, Dhikala and Bijrani zones. Subsequently, species expansion varied from medium to high in the south-western zone of the park. Other eastern and northern regions such as Morghatti, Halduparao, Mundiyapani, Hathikund and Kalagarh displayed a more recent movement of the species. In the Rajaji NP, very high infestation was seen in the southern and western regions including Haridwar, Dholkhand, Motichur and the Chilla range. Other regions such as Chillawali, Kansrao, and Mohand showed a more recent movement of the species in the northern and eastern areas (Figure 2). |
| Future Climate projections: CCCMA, CSIRO and HadCM3 climate models. Potential distribution of Lantana camara was extensive towards the north-western, eastern, and south-western regions under the CCCMA model. A slight range shift from moderate to very high invasion potential class was projected in the north-eastern and southwestern regions of the study area from the time slice 2020-2080 (Figure 3). A remarkable upward and downward expansion was also evident in response to the increasing temperatures, resulting in further spread of Lantana camara. The density of infestation by the species showed similar increasing trends in both scenarios. Thus, Lantana camara is likely to expand its invasion regime in the future as per the CCCMA model. |
| In the CSIRO model, potential distribution of Lantana camara showed altitudinal expansion under varied time slices. Greater expansion was observed towards the upward and downward regions of the study area (Figure 4). Similar to the CCCMA model, very high potential distribution was observed under both A2a and B2a emission scenarios in increasing order of magnitude from the current distribution to 2080. However, the changes in the predicted areas were more in magnitude here as compared to the CCCMA model. Under future climate projections ranging from the time slice 2020-2080, considerable range shift from moderate to very high probability of invasion potential was projected in the northern and southern regions of the study area, i.e., altitudinal shifts. This was unlike the CCCMA model where lateral shift or change in the western and eastern regions was more evident. Increase in high and moderate ranges in the central regions to very high and high suitability ranges, respectively, is indicative of a shift in the invasive regime. The projected future distribution maps for B2a scenario for 2020, 2050 and 2080 were almost similar to the A2a scenario in directional projections of invasion; however, they differed in the magnitude of infestation. |
| The HadCM3 model showed a mixed response of the potential distribution of Lantana camara with expansion along all gradients viz. lateral, upwards and downwards (Figure 5). Unlike the other two climate models, the HadCM3 model predictions revealed that A2a and B2a emission scenarios were significantly different from each other in the magnitude of infestation, with the infestation being more prominent in the former. Considerable range shift from low to very high probability of invasion potential was projected in the eastern, south-western and north-eastern regions of the study area. A remarkable southern and northern expansion was also evident, especially in the north-eastern region, in response to the increasing temperatures. The projected future distribution maps for the B2a scenario for 2020, 2050 and 2080 were almost similar to the A2a scenario in so far as the directional projections of invasion were concerned. The only difference lies in the directional spread in 2080, where only an upward movement is observed unlike A2a scenario is evident. |
| The increase in the potential distribution area was more under A2a than B2a (Table 2) across all the climate models, thus further ratifying that the species will likely expand its invasion regime under warmer climate (A2a scenario). |
| Current and future climate projections: Rajaji and Jim Corbett National Park (NP). High Lantana camara infestation was observed in Jim Corbett (Figure 6) and Rajaji NP (Figure 7) in current as well as future climate models. The species spread was more remarkable in both these NP as compared to the overall study area. The density of infestation was observed to increase with time from 2020 to 2080 for all climate models. Aligning to the common projections of greater infestation under A2a emission scenario than B2a, projections in the Jim Corbett and Rajaji NP regions were no exception across all the climate models. |
| Looking at the direction of spread, it seems that Lantana camara will consolidate its hold on the latitudes in which it currently occurs, and show lateral, upward and downward expansion in the future. All models were consensus on range shifts across time slice 2020-2080 with more infestations under A2a than under B2a scenario, indicating the preference of Lantana camara for warmer climatic conditions that are likely to be available with future climate change. Also, the change was more pronounced in the CSIRO model than the other two. |
| Analysis of variable contribution to species models |
| The environmental variables that were most influential in predicting the occurrence of Lantana camara for all the future climate models were diurnal temperature range, annual precipitation, and potential evapotranspiration of March, with variable contribution above 5% (Table 3). However, the relative contribution of these environmental variables varied among the three climate models and across time slices. |
| The diurnal temperature range was the most influential predictor variable, with its contribution ranging between 30.9-31.5% across all models in the time slice 2020 under the A2a scenario. The contribution increased in the range of 31 -34% in 2050 and 32-36% in 2080. Annual precipitation and potential evapotranspiration followed a similar trend though the contribution percentage was more pronounced under the B2a scenario. Thus, a general trend of increasing contribution of temperature-related predictor variables in the A2a model and that of precipitation-related predictor variables in the B2a model was observed. |
| In all the models, temperature and precipitation alone contributed more than 50% of the entire variable contribution. The contribution of variables like elevation, minimum temperature of coldest period and precipitation of driest period was low, but significant nonetheless. Thus, Maxent model’s internal Jackknife test of variable importance indicated that diurnal temperature, annual precipitation and potential evapotranspiration of March were the most important predictors of potential distribution of Lantana camara under future climate change. |
| Discussion |
| Projecting species distributional shifts across time |
| The potential distribution under future projections were strikingly similar to the current but were much larger in extent under all climate models and scenarios. Future predictions seem to be robust in most of the regions of the study area for all the climate models. Models suggest that most of the southern and western regions of the Western Himalaya may become infested with Lantana by 2080, because of the temperature increase. Pole ward shift of invasive species due to higher temperature and elevated precipitation has been reported in several other studies [40-42]. Given the prolific nature of Lantana camara, its ability to invade fragmented habitats, greater amplitude of ecological tolerance, and high dispersal ability and reproduction rate, the species may be able to benefit from the widening of climatically suitable areas. Although the analysis of model projections showed varying degree of range expansion, all models concurred that range expansion was greater in magnitude and direction under the A2a scenario as compared to B2a (Table 4). |
| The CSIRO and HadCM3 showed the largest modeled area under high potential distribution of Lantana camara across all time units under the A2a scenario. These were also the areas with high clamping. Clamping was considerably lower under the A2 scenarios and climate model CSIRO (<0.17). Nevertheless, clamping suggests that the results remain robust for a majority of the study area. Also, the projected potential distribution of Lantana camara is expected to expand across time slices viz. 2020, 2050 and 2080. The results further revealed that model gains and AUC values for Lantana camara were higher in HadCM3 models as compared with those of the other two models. The expansionary trend revealed by Lantana camara was in agreement with Dobhal and Kumar [30,32]. In agreement with this, the results showed that the species will have the largest expansion in its range (44%) by the year 2080. |
| Predictor variables |
| Results revealed that climatic variables were better fit to training data and accuracy of predictions. This suggests that at the meso-scale, distribution limits of Lantana camara in the Western Himalayan regions are mainly mediated by climate predictor variables viz., temperature and precipitation. Almost all physiological processes of Lantana are influenced by temperature, which in itself depends on the surrounding physical environment [43-45]. The physiological processes that are dependent on temperature include reproduction and allelopathic reactions [46], rates of dispersal [47] and ability to survive under adverse conditions [48]. Hence, distribution limits of Lantana are highly dependent on optimal temperatures as illustrated by the species potential distribution niche models. |
| Sources of errors and uncertainties in climate modeling |
| There are several sources of errors and uncertainties originating from location data and climate scenarios when predicting current and future ranges. To reduce these uncertainties associated with location data, records capturing the most well-known, long-standing and easily verifiable locations of species were used in this study. The climate change variables are based on projected changes in population, technology, socio-economic and policies [11]. These are generated through weather station data that are prone to interpolation errors and biases associated with uneven spatial and temporal distribution of the weather stations [24]. In addition, climate surfaces generated by the interpolation of weather station data may have spatial auto-correlation as closer stations are more likely to have the same climatic conditions. The uncertainty stemming from the future projections is overcome by using several models in the present study. This approach presumes that the different models are internally sound and represent at least a part of the elusive truth by combining a range of information [49]. |
| Conclusion |
| Climate change affects all components of the earth system. Yet individual components of the system react with a broad range of response times to increasing atmospheric GHG concentrations. The combined consideration of spatial and non-linear climate envelope and environmental variables applied within the context of species interaction dynamics may be useful in assessing the potential consequences of climate change for plant invasion. However, lack of empirical data, complexity of current and future projection models, and uncertainty are inherent and remain challenges to be overcome in the future. This research was undertaken to showcase the importance of modeling tools in assessing invasion potential under climate change trajectories, with special emphasis on Lantana camara in the Western Himalayan regions. |
| Three climate-change models derived from HadCM3 (Hadley Centre for Climate Prediction and Research’s General Circulation Model), CSIRO (Commonwealth Scientific and Industrial Research Organization) and CCCMA (Canadian Centre for Climate Modeling and Analysis) for the period of 2020-2080 including two scenarios A2a and B2a were used to obtain likely estimates of invasion potential distribution of Lantana camara in future. A2a scenario envisions population growth to 15 billion by the year 2100 and rather slow economic and technological development, and B2a, envisions slower population growth (10.4 billion by 2100) with a more rapidly evolving economy, more emphasis on environmental protection, lower emissions and therefore a less pronounced future warming. |
| The area of predicted future suitability is strikingly similar to current situation and much larger in extent under all predicted future scenarios. Future predictions seem to be robust in most of the regions of study area for all climate models. Models suggest that most of the southern and western regions of western Himalaya may become suitable by 2080 because of the increase in temperature. Higher temperatures and elevated precipitation regime triggered pattern is reminiscent of studies on invasive species responding to climate change by shifting their range toward the poles. However, given the prolific nature of Lantana camara, its ability to invade fragmented habitat, greater amplitude of ecological tolerance, high dispersal ability and reproduction rate, the species may be able to benefit from the widening of areas climatically suitable. The analysis of model projections showed varying degree of range expansion; however, all models were consensus on that under A2a range expansion was greater in magnitude and direction with respect of B2a scenarios. The results support the view that species ecological traits such as habitat will influence their response to changing climate causing vulnerability to native population extinction. The study such as these will henceforth enable decisions makers to develop management strategies for invasive species keeping the future in the purview. |
References
|
Tables and Figures at a glance
| Table 1 | Table 2 | Table 3 | Table 4 |
Figures at a glance
 |
 |
 |
 |
| Figure 1 | Figure 2 | Figure 3 | Figure 4 |
 |
 |
 |
| Figure 5 | Figure 6 | Figure 7 |
Relevant Topics
- Atmosphere
- Atmospheric Chemistry
- Atmospheric inversions
- Biosphere
- Chemical Oceanography
- Climate Modeling
- Crystallography
- Disaster Science
- Earth Science
- Ecology
- Environmental Degradation
- Gemology
- Geochemistry
- Geochronology
- Geomicrobiology
- Geomorphology
- Geosciences
- Geostatistics
- Glaciology
- Microplastic Pollution
- Mineralogy
- Soil Erosion and Land Degradation
Recommended Journals
Article Tools
Article Usage
- Total views: 16192
- [From(publication date):
December-2013 - Dec 09, 2025] - Breakdown by view type
- HTML page views : 11164
- PDF downloads : 5028